 2015-02-14
2015-02-14 2728
2728Характерные для каждого вида нуклеиновых кислот мононуклеотиды, объединяясь в количестве нескольких сотен, а иногда и тысяч, в единую молекулу, образуют громадные полимерные цепи. Таким образом по своему химическому строению нуклеиновые кислоты являются полирибонуклеотидами (РНК) и полидезоксирибонуклеотидами (ДНК). Соединение нуклеотидных остатков в молекулах нуклеиновых кислот осуществляется одним и тем же путём, с помощью фосфодиэфирных связей, образуемых между парами нуклеотидов остатками фосфорной кислоты, которые связывают 3¢- и 5¢-атомы соседних нуклеотидов.
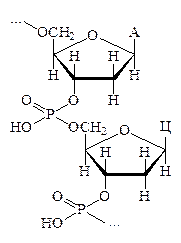
Наиболее высокими молекулярными массами обладают молекулы ДНК, для которых типичной величиной является масса от 5 до 10 млн. дальтон. Это самые большие молекулы в природе. Молекулярные массы РНК изменяются в достаточно широких пределах. При этом выделяют низкомолекулярные РНК – т-РНК (18–35 тыс. дальтон) и высокомолекулярные РНК – р-РНК (600 тыс. – 1 млн. 200 тыс. дальтон) и и-РНК (1 млн. – 4 млн. дальтон).
Первичная структура ДНК
Представляет собой изогнутую спиральную цепь соединенных ковалентными связями нуклеотидов.
Установление первичной структуры ДНК сложнее установления первичной структуры белка. Это связано с однообразием её строения. В 1977 году была определена полная нуклеотидная последовательность ДНК бактериофага YC 174 (пси-икс 174). Это выдающееся достижение ознаменовало собой начало новой эры в биохимии генов и хромосом. С тех пор была расшифрована нуклеотидная последовательность целого ряда генов. В настоящее время в принципе можно определить последовательность нуклеотидов любой ДНК.
Решающий успех в установлении первичной структуры ДНК был достигнут благодаря 3-м основным достижениям.
Первым из них явилось открытие рестриктирующих эндонуклеаз, которые расщепляют молекулы ДНК только в сравнительно небольшом числе специфических точек. Это позволило расщеплять молекулу ДНК на определённые фрагменты разными способами с образованием перекрывающихся последовательностей.
Вторым важным достижением оказалось усовершенствование электрофоретических методов разделения фрагментов ДНК в соответствии с числом содержащихся в них нуклеотидных остатков (с общим числом приблизительно 200 нуклеотидов, различающихся всего на один остаток).
Третьим достижением стала разработка методов клонирования ДНК, которое сделало возможным получение достаточно большого количества чистых модельных генов – исходного материала для определения первичной структуры ДНК.
На основании всего этого было предложено 2 принципиальных подхода к определению первичной структуры нуклеиновых кислот – метод обрыва цепи Ф. Сенгера и Р. Коулсона и метод химического расщепления (А. Максама и В. Гильберта).
Метод обрыва цепи основан на получении полного набора убывающих по числу нуклеотидных остатков копий одноцепочечной ДНК с помощью ДНК-полимеразной реакции. При этом используются 4 реакционные смеси, в каждую из которых добавляют, помимо 4-х мономеров, аналог одного из оснований (2¢,3¢-дидезоксианалог) в качестве терминатора роста цепи. Затем цепи в каждой смеси разделяют с помощью гель-электрофореза и проводят радиоавтографию с помощью радиоактивного фосфора (Р-32). По числу наблюдаемых полос устанавливают число нуклеотидных остатков ДНК или её фрагмента и выявляют положение азотистых оснований с помощью особых реакций на 3¢-конце каждого из фрагментов.
Метод химического расщепления. В соответствии с этим методом проводят химическую модификацию азотистых оснований в ДНК (реакциями алкилирования, ацилирования). Затем избирательно расщепляют цепочку ДНК по месту модифицированных оснований, проводят разделение продуктов расщепления в полиакриламидном геле с помощью электрофореза. После этого анализируют полученные электрофореграммы и по положению модифицированного азотистого основания восстанавливают первичную структуру в анализируемом фрагменте ДНК.
Недавно с использованием последних версий описанных методов была установлена полная первичная структура генома человека.
Вторичная структура ДНК
Вторичная структура ДНК представляет собой двуспиральную макромолекулу, составленную парами взаимозакрученных полидезоксирибонуклеотидных цепей, каждой из которых свойственно специфическое, но противоположное чередование нуклеотидных остатков, соединённых по принципу комплементарности.
Эта модель была предложена в 1953 г. Дж. Уотсоном и Ф. Криком на основании данных рентгеноструктурного анализа ДНК. При этом было установлено, что ДНК склонна к полиморфизму и в различных условиях (как правило, в среде с различным ионным составом) существует в виде по-разному упорядоченных волокнисто-кристаллических структур. Их существует более десятка, но наиболее распространены и изучены методом рентгеноструктурного анализа только четыре: А-, В-, С- и Т-формы ДНК. Все они являются правозакрученными спиралями. Существование такого количества вторичных структур ДНК обусловлено тем, что в каждый момент своего существования ДНК образует наиболее удобных для выполняемых ею функций конфигурации:
– А-форма молекул наиболее удобна для процесса транскрипции;
– В-форма – для репликационных процессов;
– С-форма – для образования молекулой ДНК и белками хроматина.
В 70-х годах XX века были получены данные о существовании двух новых конформаций вторичной структуры ДНК – Z-формы и SBS-конформации. Z-форма представлена левозакрученными полидезоксирибонуклеотидными цепями с зигзагообразным расположением фосфатных групп. В Z-форме ДНК участвует в различных метаболических процессах. SBS-конформация ДНК характеризуется отсутствием взаимозакрученных полидезоксирибонуклеотидных цепей в биспиральную молекулу; они располагаются бок о бок, очень легко распариваются, расходятся, что играет большую роль при биосинтезе ДНК.
Образование описанных структур ДНК (кроме SBS-конформации) возможно за счёт двух причин:
а) благодаря водородным связям между комплементарными азотистыми основаниями, обращёнными внутрь двойной спирали;
б) за счёт силы гидрофобных взаимодействий между азотистыми основаниями, собранными в «стопку» вдоль молекулы ДНК, так называемых стэкинг-взаимодействий.
Считают, что гиброфобные взаимодействия играют определяющую роль в формировании вторичной структуры ДНК, а водородные связи – всего лишь направляющую.
Биспиральные структуры в молекулах ДНК возникают не только при взаимодействии двух комплементарных полидезоксирибонуклеотидных цепей, но и в пределах одной и той же цепи. Это случается тогда, когда в комплементарных цепях ДНК присутствуют палиндромные участки. Палиндромы спирализуются сами на себя, образуя так называемые «шпильки» – структуры, приспособленные для узнавания структур ДНК ферментами и регуляторными белками. Также палиндромы служат для формирования элементов третичной структуры ДНК.
Третичная структура ДНК
Представляет собой расположение полинуклеотидных цепей в трёхмерном пространстве. Молекулы ДНК при этом подвергаются дополнительной спирализации и суперспирализации и существуют в виде линейных и кольцевых форм. В линейной форме находится большинство природных ДНК, а ДНК большинства вирусов, фагов, клеточных органелл – хлоропластов, митохондрий, центриолей, бактериальных плазмид находится в кольцевой форме. Между линейными и кольцевыми формами существуют динамические переходы:
В хромосомах эукариот ДНК находится в суперспирализованном состоянии в связи с белками, образуя дезоксирибонуклеопротеиды. В нуклеопротеидах белковый компонент представлен белками протеинами основного типа – протаминами и гистонами. Связь между белками и нуклеиновыми кислотами осуществляется посредством электростатического взаимодействия между ионизированными остатками фосфорной кислоты нуклеотидов и ионизированными радикалами лизина, аргинина и гистидина белков:
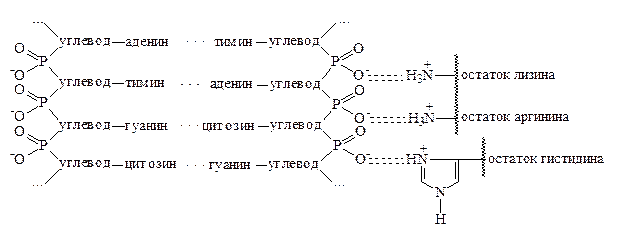
При соединении ДНК с белками происходит взаимная стабилизация и белков, и нуклеиновых кислот вследствие своеобразной реакции нейтрализации.
ДНК в соединении с белками образует элементарную единицу структуры хроматина – нуклеосому. Особенностью процесса формирования третичной структуры ДНК является также и то, что переход ДНК в суперспирализованное состояние и обратно осуществляется посредством особой группы ферментов – топоизомераз, изменяющих пространственную структуру нуклеопротеидного комплекса.
Молекулярная структура ДНК обуславливает наличие у этой нуклеиновой кислоты целого комплекса свойств, в чём-то совпадающих с общими свойствами нуклеиновых кислот, и чем-то различающихся с ними.








