 2015-02-15
2015-02-15 2654
2654Нервная регуляция сердечно-сосудистой системы является мощным инструментом изменения кровообращения. Прямой нервный контроль гладкомышечных клеток сосудов является высшим по сравнению с влиянием на сосуды гуморальных веществ, содержащихся в крови, поскольку он обеспечивает быструю и, если необходимо, локализованную регуляцию. Имеются указания, что гормональные влияния, например, надпочечников, в норме слабее, чем прямое действие симпатических нервов на сердечно-сосудистые эффекторы. С другой стороны, при конкурировании с химическими вазодилататор-ными метаболитами, образующимися при деятельности органов, ва-зоконстрикторные волокна могут уступать в своем действии на сосуды.
Из сказанного следует, что нормальное функционирование системы кровообращения в условиях постоянно меняющихся внешних воздействий осуществляется только при участии механизмов регуляции (управления, контроля). Под регуляцией физиологических систем понимают механизмы, проявляющиеся в ответ на сдвиг физиологической функции и обычно направленные на компенсацию развивающихся эффектов этих воздействий. Термин "регуляция" употребляется только в биологической и физиологической литературе.' В технических науках и в междисциплинарных изложениях ему соответствует понятие "управление" и "регулирование".
Теория автоматического регулирования предусматривает две системы регулирования процесса: по отклонению и по возмущению. В первом случае, речь идет о том, чтобы регулируемая величина всегда была как можно ближе к заданному воздействию, т.е. разность этих сигналов — отклонение или рассогласование — была бы как можно меньше, несмотря на действие внешних возмущений. Во втором случае, имеют в виду, что в системе имеется прямая связь от внешнего воздействия, называемого возмущением, через регулирующее устройство к регулируемой величине.
Примером регуляции по отклонению могут быть механизмы нервной регуляции постоянства артериального давления. В этом случае "объектом управления" являются сердце и сосуды, так как даже небольшие изменения их деятельности легко модулируют уровень артериального давления. "Управляющим устройством" служат центральные структуры, связанные с поддержанием нейрогенного сосудистого тонуса и нервных влияний на сердце. В этом "управляющем устройстве" выделяют "детектор ошибки", определяющий разность между входным сигналом и задающим, и "регулятор" — бульбарный вазомоторный центр, который на входе воспринимает "сигнал ошибки", а управляющий сигнал ориентирует на "объект управления".
326
Примером регуляции артериального давления по возмущению могут служить барорецепторные рефлексы артериального русла, которые сводят к минимуму колебания системного давления при различных воздействиях (возмущениях) на организм.
Непременным условием рассматриваемого вопроса является цель регуляции (управления). В физиологии принято употреблять выражение не "цель", а "физиологическое значение (роль)" регуляторного механизма. Основной целью функционирования кровообращения, т.е. важнейшей физиологической ролью (значением) кровообращения является своевременная доставка кислорода, питательных и других веществ, а также жидкости всем клеткам тела, отведение накапливаемых метаболитов и тепла из тканей, и этим — поддержание состава внутренней среды вокруг клеток, необходимого для осуществления их метаболизма и функции.
Две части системы управления — управляющее устройство и объект управления образуют вместе контур управления. В кровообращении в качестве объекта управления можно выделить капиллярное русло и течение крови по нему, обеспечивающие питание тканей, тогда управляющими устройствами являются отделы сердечнососудистой системы и свойства крови, обеспечивающие необходимый кровоток в капиллярном русле в ходе быстротекущих событий. Локализация управляющего устройства, в целом, устанавливается достаточно четко, особенно, если речь идет о центральной нервной регуляции.
Спинальный уровень регуляции. Эффекторными клетками, осуществляющими регуляцию сердца и сосудов на уровне спинного мозга, являются симпатические преганглионарные нейроны. Они обладают спонтанной фоновой импульсной активностью, частота которой коррелирует с изменениями частоты сокращений сердца и колебаний артериального давления. Сложная организация межнейронных связей между симпатическими преганглионарными нейронами свидетельствует о том, что ядра боковых рогов спинного мозга могут обеспечивать преобразования нервных импульсов по дивергентному и конвергентному типу. Таким образом, спинальные нейроны представляют собой важное промежуточное звено по пути нисходящих нейрогенных влияний, адресованных эффекторам в сердечно-сосудистой системе.
Особо стоит вопрос о рефлекторной функции спинного мозга в регуляции кровообращения. Перерезка в эксперименте спинного мозга на уровне шейных и верхних грудных сегментов приводит к значительному снижению (до 60 мм рт.ст.) артериального давления. Однако, если перерезка выполнена в щадящих условиях препаровки (например, при помощи ультразвукового ножа), уже через несколько часов давление восстанавливается практически до нормальных величин. В этих же условиях восстанавливаются и сосудодвигательные рефлексы, возникающие в ответ на раздражение соматических нервов, рецепторов мышц и кожи, механорецепторов брюшины и рецепторов растяжения сердечных камер. Эти факты свидетельствуют,
327
что спинной мозг при определенных условиях может являться уровнем замыкания рефлексов сердечно-сосудистой системы. В то же время в естественных условиях рефлексы сердечно-сосудистой системы осуществляются при обязательном участии бульбарных и ги-поталамических образований головного мозга.
Бульбарный уровень регуляции. Современная концепция центральной регуляции кровообращения исходит из возможности существования серии параллельных контуров регуляции, характеризующихся различными уровнями замыкания рефлекторных дуг. Высокая степень автономности этих контуров регуляции не исключает тесного взаимодействия между ними, причем, именно это взаимодействие определяет конечный результат в виде детерминированных, приспособительных реакций сердца и сосудов в ответ на разнообразные внешние и внутренние раздражители.
Современные представления о бульбарных механизмах регуляции кровообращения основаны на исследованиях, в которых показано влияние как перерезок на разных уровнях, так и стимуляции различных бульбарных структур на артериальное давление и частоту сердечных сокращений, а также на импульсации в симпатических и парасимпатических нервах. Раздражение продолговатого мозга и вар-олиева моста оказывает либо прессорные, либо депрессорные изменения АД в зависимости от локализации раздражающих электродов: прессорные зоны располагаются, как правило, ростральнее и лате-ральнее депрессорных. При перерезке на уровне ядер лицевого нерва отделяется расположенная ростральнее часть прессорной области, тогда как депрессорная остается интактной. В этом случае снижается артериальное давление и уменьшается импульсация в эфферентных вегетативных нервах. Перерезка продолговатого мозга на уровне задвижки ведет к еще большему снижению давления и полному прекращению импульсации по вегетативным нервам. Бульбар-ная медиальная депрессорная область оказывает угнетающее тоническое влияние на спинальные преганглионарные нейроны, способные к спонтанной импульсной активности.
Таким образом, латеральные участки бульбарной ретикулярной формации содержат скопления спонтанно активных нейронов, которые через нисходящие пути спинного мозга оказывают стимулирующее влияние на спинальные преганглионарные симпатические нейроны. Эти нейроны образуют латеральную "прессорную" область продолговатого мозга. Функции этих нейронов осуществляются в реци-прокном взаимодействии с нейронами медиальной 'депрессорной" зоны.
Указанный медуллярный комплекс, реализующий влияния на сердечно-сосудистую систему через симпатические нервы, называют бульварным сосудодвигательным центром. Функционально он включает в себя и структуры дорзального ядра блуждающего нерва, обеспечивающие тонические кардиоингибиторные влияния. Поэтому, говорят о едином бульбарном центре сердечно-сосудистой системы, получающем информацию от различных рецепторных зон и обеспечивающем поддержание циркуляторного гомеостаза. Это достигается, бла-
годаря реципрокному взаимодействию симпатических и парасимпатических рефлекторных влияний на сердце, а также дозировкой тонических констрикторных влияний симпатических нервов на сосуды.
В вентролатеральных отделах продолговатого мозга сосредоточены образования, соответствующие по своим характеристикам тем представлениям, которые вкладывают в понятие "вазомоторный центр". Здесь сконцентрированы нервные элементы, играющие ключевую роль в тонической и рефлекторной регуляции кровообращения. Активация структур ростральной части вентральных отделов продолговатого мозга увеличивает параметры резистивной функции сосудов, уменьшает емкость органного сосудистого русла и ослабляет его обменную функцию. Возбуждение структур каудальной части вентральных отделов продолговатого мозга ведет к противоположному эффекту указанных сосудистых функций. Нейроны ростральной части вентральных отделов продолговатого мозга являются звеном дуг собственных и сопряженных сосудистых рефлексов, а нейроны каудальной части — модулируют реализацию на сосуды указанных рефлексов.
Сердечно-сосудистый центр продолговатого мозга оказывает тоническое возбуждающее влияние на преганглионарные симпатические нейроны тораколюмбального отдела спинного мозга. Однако, его влияния являются не единственными связями, конвергирующими на спинальных вегетативных нейронах. Считается, что в медуллярной области происходит переключение нисходящих возбуждающих и тормозных кардиоваскулярных влияний, исходящих из вышележащих отделов центральной нервной системы. С другой стороны, собственная тоническая активность спинальных нейронов в норме до такой степени зависит от нисходящих бульбарных и супрабульбарных влияний, что эти нисходящие влияния полностью доминируют над их спонтанной активностью. Несмотря на то, что сегментарные спи-нальные вазомоторные рефлексы (например, побледнение кожи над областью внутрибрюшинной воспалительной реакции) могут иметь место, замыкание собственных сердечно-сосудистых рефлексов происходит не ниже, чем на бульбарном уровне. Однако и бульбарный уровень регуляции в естественных условиях модулируется сложной совокупностью нисходящих к нему влияний, среди которых ведущую роль играют нейро-гуморальные влияния гипоталамического происхождения.
Гипоталамические влияния. При электрическом или химическом раздражении гипоталамических структур имеют место изменения артериального давления, причем раздражение одной и той же структуры может вызывать как прессорные, так и депрессорные сдвиги. Несмотря на преобладание прессорных элементов и их диффузное распределение по гипоталамусу, концентрация депрессорных элементов в переднем гипоталамусе существенно выше, чем в заднем. Характер реакции на раздражение гипоталамуса зависит не только от места раздражения, но и от его параметров, а также от функционального состояния сердечно-сосудистой системы. Наиболее
329
универсальным механизмом реакции сердечно-сосудистой системы на возбуждение гипоталамуса является активация симпатической системы. Типичным результатом возбуждения гипоталамических структур в этом случае является альфа-адренергическая констрикция периферических сосудов в сочетании с положительными хронотроп-ными и инотропными влияниями на сердце.
В то же время симпатические эффекторы гипоталамических влияний могут обусловить и дилатацию периферических сосудов, причем, существует несколько механизмов реализации такого рода влияний. Это может быть, например, симпатическая холинергическая или бета-адренергическая дилатация, а также результат центрального нисходящего гипоталамического торможения бульбарных сердечно-сосудистых центров.
Гипоталамические влияния на кровообращение могут реализовы-ваться не только рефлекторным, но и гуморальным путем. Гипоталамус вместе с гипофизом образуют систему, регулирующую работу большинства эндокринных желез, в том числе и тех, которые участвуют в регуляции кровообращения. Возбуждение гипоталамуса сопровождается выделением вазопрессина в гипофизе и норадреналина в надпочечниках с их последующим действием на сердце и сосуды, а также выделение ренина почками в результате прямых нейроген-ных влияний на их юкстагломерулярный аппарат. Конечным результатом этого процесса является повышение артериального давления за счет образования и появления в крови ангиотензина II.
Существенной особенностью гипоталамической регуляции кровообращения является то, что этот отдел центральной нервной системы оказывает дифференцированные влияния на сердце и некоторые сосудистые бассейны. Так, раздражение гипоталамических центров "защитных" реакций организма вызывает увеличение сердечного выброса, повышение артериального давления, сужение сосудов внутренних органов, но при этом имеет место расширение артериальных сосудов скелетных мышц. Стимуляция пищевых центров в латеральном гипоталамусе приводит, наоборот, к расширению сосудов желудочно-кишечного тракта и их сужению в скелетных мышцах.
Гипоталамические нейроны получают сигналы практических от всех экстеро- и интероцепторов организма, включая артериальные бароре-цепторы, импульсы от которых поступают в передний гипоталамус. Кроме того, гипоталамические нейроны получают информацию об изменениях внутренней среды организма (температуры, кислотности, осмолярности и др.). Следовательно, существует еще одна рефлекторная функция гипоталамуса, физиологический смысл которой заключается в обработке и интеграции этой информации, и включения в общую реакцию сердечно-сосудистой системы, направленную на поддержание гомеостазиса. Эта функция гипоталамуса включает в себя и контроль деятельности ниже лежащих бульбарных и спинальных структур, ответственных за регуляцию кровообращения.
Важная роль гипоталамуса заключается также в координации кровообращения с другими висцеральными и соматическими функциями. В частности, известно, что некоторым эмоциональным состоя-
330
ниям, связанным с активацией гипоталамических структур, соответствуют и определенные изменения гемодинамики. Поэтому считается, что гипоталамус осуществляет координацию соматомоторных и вегетативных проявление эмоционального поведения.
Кортикальные влияния. Хотя изучению роли коры больших полушарий в регуляции кровообращения уделялось пристальное внимание, результаты таких исследований до настоящего времени весьма противоречивы. В экспериментах на животных показано, что удаление обоих полушарий не вызывает каких- либо существенных • изменений кровообращения. В то же время электрическая стимуляция участков лобной и теменной областей коры ведет к изменениям артериального давления в результате сужения или расширения сосудов, причем, в зависимости от параметров раздражения стимуляция одних и тех же точек может вызвать как вазоконстрикцию, так и вазодилатацию.
Если наличие коркового компонента показано в реализации без-условнорефлекторных реакций кровеносных сосудов, то влияние коры больших полушарий на сердце связано с высшей нервной деятельностью — реализацией эмоциональных, поведенческих реакций и условных рефлексов. Последние, по-видимому, могут образовываться лишь на основе сопряженных кардиальных рефлексов. Изменения кровообращения, обусловленные собственными рефлексами сердечно-сосудистой системы, воспроизвести условнорефлекторным путем не удалось. Условнорефлекторные реакции сердца являются основной частью сложных поведенческих рефлексов. У человека эти реакции могут быть вызваны и словесными раздражителями (речевыми сигналами). Однако, произвольно изменить параметры сердечной деятельности можно только косвенным путем, через заведомо управляемые системы: дыхание или скелетную мускулатуру, а также в некоторых случаях произвольно — изменяя эмоциональное состояние организма.
Важное значение имеют кортикальные механизмы в реализации изменений кровообращения при стрессовых и невротических состояниях человека. В исследовании этих вопросов наиболее продуктивным является сочетание физиологических и психологических методов.
Общая схема центральной регуляции. На рис.7.27 представлена общая схема организации центральных звеньев нервной регуляции кровообращения. Сердечно-сосудистый центр продолговатого мозга состоит из двух зон (А и Б), одна из которых определяет сужение сосудов (А'), ускорение и усиление сердцебиений (А"), тогда как другая — расширение сосудов (БI) и замедление частоты сокращений сердца (БII). Тонически активные нейроны зоны А непосредственно или через промежуточные нейроны возбуждает прегангли-онарные и ганглионарные симпатические нейроны, иннервирующие сердце и сосуды, и в то же время тормозят нейроны зоны Б. Последние активируются сигналами барорецепторов сердечно-сосудистой системы. Через короткие внутрицентральные пути нейроны
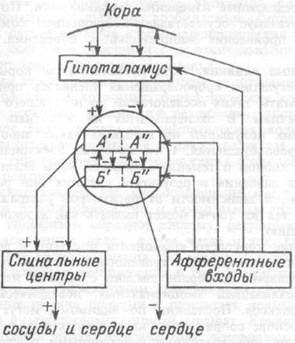
Рис.7.27. Схема организации центральных звеньев нервной регуляции сердечно-сосудистой системы. Объяснение в тексте.
зоны Б тормозят нейроны зоны А, а посредством нисходящих тормозных путей — и преганглионарные симпатические нейроны. Кроме того, через моторное ядро блуждающих нервов нейроны зоны Б влияют на ганглионарные парасимпатические нейроны сердца. В результате обе зоны (А и Б), получая основную сигнализацию от рецепторов сердечно-сосудистой системы, являются основой поддержания кровообращения, сочетательно подвергая сердце тоническому контролю тормозных вагусных и возбуждающих симпатических волокон, тогда как постоянный контроль сосудов обеспечивается только симпатическими вазоконстрикторными волокнами.
Афферентная нервная система является источником сигналов собственных и сопряженных рефлексов. Вход первых ограничен продолговатым мозгом, причем барорецепторы сердечно-сосудистой системы связаны с зоной Б, а хеморецепторы — с зоной А. С последней связаны также и некоторые афференты сопряженных рефлексов (от рецепторов кожи, мышц и внутренних органов), хотя организация этих связей известна лишь в общих чертах.
Поскольку сердечно- сосудистый центр продолговатого мозга регулирует основные параметры системной гемодинамики, даже будучи
отключенным от связей с вышележащими нервными структурами, он функционирует как истинный "центр", обладающий значительной независимостью. Следовательно, супрабульбарные структуры признаются несущественными ни для прямой тонической регуляции кровообращения, ни для осуществления сердечно-сосудистых рефлексов. В то же время при общих реакциях организма (поведенческих, эмоциональных, стрессорных и др.) кора мозга, гипоталамус и другие супрабульбарные структуры могут вмешиваться в деятельность сердечно-сосудистой системы, изменяя путем нисходящих (модулирующих) влияний возбудимость бульбарного центра.
Вазомоторные центры спинного мозга самостоятельного значения в регуляции кровообращения не имеют. Действующими элементами спинального уровня являются лишь симпатические преганглионар-ные нейроны, которые входят в состав эфферентного звена буль-барных рефлекторных дуг. Только в случае травм, приводящих к утрате связей продолговатого мозга со спинным, последний, спустя некоторое время, приобретает способность осуществлять вазомоторные рефлексы на импульсы спинномозговых афферентов.

