 2015-03-27
2015-03-27 2471
2471Эндокринная система занимает одно из центральных мест в управлении различными процессами жизнедеятельности на уровне целого организма. Эта система с помощью продуцируемых гормонов непосредственно участвует в управлении метаболизмом, физиологией и морфологией различных клеток, тканей и органов (см. приложение 5).
Гормоны— это биологические высокоактивные вещества, образующиеся в железах внутренней секреции, поступающие в кровь и оказывающие регулирующее влияние на функции удаленных от места их секреции органов и систем организма.
Гормоны определяют интенсивность синтеза белка, размеры клеток, их способность делиться, рост всего организма и его отдельных частей, формирование пола и размножение; различные формы адаптации и поддержание гомеостаза; высшую нервную деятельность.
Принцип физиологического действия гормонов состоит в том, что они, попадая в кровяное русло, разносятся по всему организму. Гормоны оказывают свое физиологическое действие в минимальных дозах. Например, 1 г адреналина может активировать работу 100 млн изолированных сердец. На мембранах клеток имеются рецепторы к многим гормонам. Молекула каждого типа гормона может
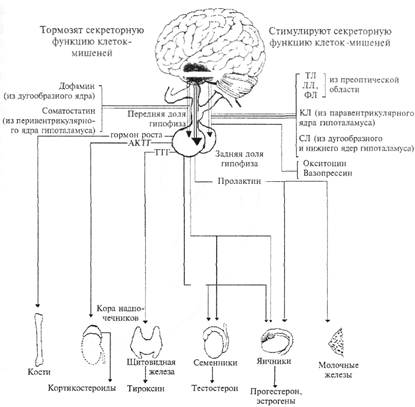
(Рис. 4.6. Схема нейроэндокринной системы. АКТГ — адренокортикотропный гормон (кортикотропин); КЛ — кор-тиколиберин; ЛГ — лютеинизирующий гормон; ЛЛ — люлиберин; СЛ — соматолиберин; ТЛ — тиреолиберин; ТТГ — тиреотропный гормон; ФЛ — фоллиберин; ФСГ — фолликулостимулирующий гормон.
соединиться только со «своим» рецептором на клеточной мембране (принцип: молекула гормона подходит к рецептору, как «ключ к замку»). Такие клетки называют клетками-мишенями. Например, для половых гормонов клетками-мишенями будут клетки половых желез, а для адренокортикотропного гормона (АКТГ), который выбрасывается при стрессе, клетками-мишенями будут клетки коры надпочечников. Несколько примеров взаимоотношения между гормонами гипофиза и органами-мишенями показано на рис. 4.6. Нарушение того или иного звена эндокринной системы
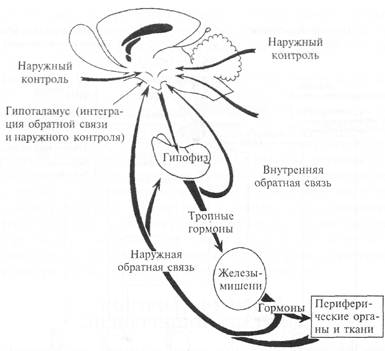
Рис. 4.7. Общая схема регуляции эндокринной системы организма по принципу «обратной связи».
может значительно изменить нормальное течение физиологических процессов, приводя к глубокой патологии, часто несовместимой с жизнью.
Между нервной и эндокринной системами имеет место теснейшая функциональная взаимозависимость, которая обеспечивается различными видами связей (рис. 4.7).
ЦНС оказывает влияние на эндокринную систему двумя путями: с помощью вегетативной (симпатической и парасимпатической) иннервации и изменения активности специализированных нейроэндокринных центров. Проиллюстрируем это важное положение на примере поддержания уровня глюкозы в крови при резком снижении концентрации глюкозы в кровяной плазме (гипогликемия). Поскольку глюкоза абсолютно необходима для функционирования головного мозга, гипогликемия не может продолжаться долго. Эндокринные клетки поджелудочной железы отвечают на гипогликемию секрецией гормона глюкагона, который стимулирует выделение глюкозы из печени. Другие эндокринные клетки поджелудочной железы отвечают на гипогликемию, напротив. снижением выделения другого гормона — инсулина, что приводит к снижению утилизации глюкозы всеми тканями, за исключением головного мозга. Глюкорецепторы гипоталамуса реагируют на гипогликемию, усиливая освобождение глюкозы из печени через активацию симпатической нервной системы. Кроме этого, активируется мозговой слой надпочечников и выбрасывается адреналин, который снижает утилизацию глюкозы тканями организма, а также способствует освобождению глюкозы из печени. Другие нейроны гипоталамуса реагируют на гипогликемию, стимулируя выделение из коры надпочечников гормона кортизола, который усиливает синтез глюкозы в печени, когда это депо истощается. Кортизол также тормозит инсулинактивируемую утилизацию глюкозы всеми тканями, за исключением головного мозга. Результатом совместных реакций нервной и эндокринной систем является возвращение к норме концентрации глюкозы в кровяной плазме в течение 60-90 мин.
В определенных условиях одно и то же вещество может выполнять роль гормона и медиатора, а механизм в обоих случаях сводится к специфическому взаимодействию молекулы с рецептором клетки-мишени. Сигналы от эндокринных желез, роль которых выполняют гормоны, воспринимаются специализированными нервными структурами и в конечном итоге трансформируются в изменение поведения организма и в ответы эндокринной системы. Последние становятся частью регуляторных реакций, образующих нейроэндокринную интеграцию. На рис. 4.7 показаны возможные виды взаимоотношений нервной и эндокринной систем. В любом конкретном случае реально используются лишь некоторые из этих путей.
Гипофиз, нижняя мозговая железа, — сложный эндокринный орган, расположенный в основании черепа в турецком седле основной кости, анатомически связан ножкой с гипоталамусом. Он состоит из трех долей: передней, средней и задней. Передняя и средняя доли объединяются под названием аденогипофиз, а задняя доля называется нейрогипофизом. В ней рогипофизе выделяют два отдела: передний нейрогипофиз, или срединное возвышение, и задний нейрогипофиз, или заднюю долю гипофиза.
Гипофиз содержит очень развитую сеть капилляров, стенки которых имеют специальное строение, так называемый фенестрированный
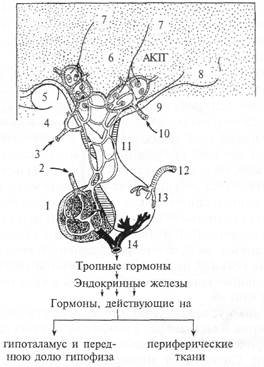
Рис. 4.8. Схема гипоталамо-гипофизарной сети капилляров.
1 — передняя доля гипофиза; 2 — артерия передней доли гипофиза; 3 — гормональная обратная связь; 4 — артерия переднего гипоталамуса; 5 — зрительный перекрест; 6 — тиреотропный освобождающий гормон; 7 — аксон; 8 — мамиллярное тело; 9 — гипоталамическая артерия; 10 — гормональная обратная связь; II — воротная система; 12 — артерия задней доли гипофиза; 13 — задняя доля гипофиза; 14— венозный отток; АКТГ — адренокортикотропный гормон.
(продырявленный) эпителий. Эту сеть капилляров называют «чудесной капиллярной сетью» (рис. 4.8). На стенках капилляров оканчиваются синапсами аксоны нейронов гипоталамуса. Благодаря этому нейроны выбрасывают из синапсов на стенках этих сосудов синтезированные белковые молекулы непосредственно в кровяное русло. Все нейрогормоны представляют собой гидрофильные соединения, для которых на поверхности мембраны клеток-мишеней имеются соответствующие рецепторы. На первом этапе происходит взаимодействие нейрогормона с соответствующим рецептором мембраны. Дальнейшая передача сигнала осуществляется внутриклеточными вторичными посредниками. Схема нейроэкдокринной системы организма человека представлена в приложении 5.
Контроль секреции задней доли гипофиза. Задняя доля, или нейрогипофиз, — эндокринный орган, аккумулирующий и секретирующий два гормона, синтезируемые в крупноклеточных ядрах переднего гипоталамуса (паравентрикулярном и супраоптическом), которые затем транспортируются по аксонам в заднюю долю. К нейрогипофизарным гормонам у млекопитающих относятся вазопрес-син, или антидиуретический гормон, регулирующий водный обмен, и окситонин, гормон, участвующий в родовом акте.
Под влиянием вазопрессина увеличивается проницаемость собирательных трубок почки и тонус артериол. Вазопрессин в некоторых синапсах нейронов гипоталамуса выполняет медиаторную функцию. Его поступление в общий кровоток происходит в случае увеличения осмотического давления плазмы крови, в результате активируются осморецепторы — нейроны супраоптического ядра и околоядерной зоны гипоталамуса. При снижении осмолярности плазмы крови активность осморецепторов тормозится и секреция вазопрессина уменьшается. С помощью описанного нейроэндокринного взаимодействия, включающего чувствительный механизм обратной связи, регулируется постоянство осмотического давления плазмы крови. При нарушении синтеза, транспортировки, выделения или действия вазопрессина развивается несахарный диабет. Ведущие симптомы этого заболевания — выделение большого количества мочи с низкой относительной плотностью (полиурия) и постоянное чувство жажды. У больных диурез достигает в сутки 15-20 л, что не менее чем в 10 раз выше нормы. При ограничении приема воды у больных наступает обезвоживание организма. Секрецию вазопрессина стимулируют уменьшение объема экстраклеточной жидкости, боль, некоторые эмоции, стресс, а также ряд препаратов — кофеин, морфин, барбитураты и др. Алкоголь и увеличение объема экстраклеточной жидкости снижают выделение гормона. Действие вазопрессина кратковременно, поскольку он быстро разрушается в печени и почках.
Окситоцин — гормон, регулирующий родовой акт и секрецию молока молочными железами. Чувствительность к окситоцину повышается при введении женских половых гормонов. Максимальная чувствительность матки к окситоцину отмечается во время овуляции и накануне ролов. В эти периоды происходит наибольшее выделение гормона. Опускание плода по родовому каналу стимулирует соответствующие рецепторы, и афферентация поступает в паравентрикулярные ядра гипоталамуса, которые повышают секрецию окситоцина. Во время полового акта секреция гормона увеличивает частоту и амплитуду сокращений матки, облегчая транспорт спермы в яйцеводы. Окситоцин стимулирует молокоотдачу, вызывая сокращение миоэпителиальных клеток, выстилающих протоки молочной железы. В результате повышения давления в альвеолах молоко выжимается в большие протоки и легко выделяется через соски. При раздражении тактильных рецепторов молочных желез импульсы направляются к нейронам паравентрикулярного ядра гипоталамуса и вызывают освобождение окситоцина из нейрогипофиза. Действие окситоцина на молокоотдачу проявляется через 30—90 с после начала стимуляции сосков.
Контроль секреции передней доли гипофиза. Большая часть гормонов передней доли гипофиза выполняет роль специфических регуляторов других эндокринных желез, это так называемые «тропные» гормоны гипофиза.
Адренокортикотропный гормон (АКТГ) — главный стимулятор коры надпочечников. Этот гормон выделяется при стрессе, разносится по кровяному руслу и достигает клеток-мишеней коры надпочечников. Под его действием из коры надпочечников в кровь выбрасываются катехоламины (адреналин и норадреналин), которые оказывают на организм симпатическое действие (подробнее этот эффект был описан выше). Лютеинизирующий гормон является главным регулятором биосинтеза половых гормонов в мужских и женских гонадах, а также стимулятором роста и созревания фолликулов, овуляции, образования и функционирования желтого тела в яичниках. Фолликулостимулирующий гормон повышает чувствительность фолликулы к действию лютеинизирующего гормона, а также стимулирует сперматогенез. Тиреотропный гормон — главный регулятор биосинтеза и секреции гормонов щитовидной железы. К группе тропных гормонов относится гормон роста, или соматотропин, — важнейший регулятор роста организма и синтеза белка в клетках; участвует также в образовании глюкозы и распаде жиров; часть гормональных эффектов опосредуется через усиление печенью секреции соматомедина (фактора роста I).
Помимо тропных гормонов, в передней доле образуются гормоны, выполняющие самостоятельную функцию, аналогичную функциям гормонов других желез. К таким гормонам относятся: пролактин, или лактогенный гормон, регулирующий лактацию {образование молока) у женщины, дифференцировку различных тканей, ростовые и обменные процессы, инстинкты выхаживания потомства у представителей различных классов позвоночных. Ли-потропины ~ регуляторы жирового обмена.
Функционирование всех отделов гипофиза тесно связано с гипоталамусом. Гипоталамус и гипофиз образуют единый структурно-функциональный комплекс, который часто называют «эндокринным мозгом».
Эпифиз, или верхняя шишковидная железа, входит в состав эпиталамуса. В эпифизе образуется гормон мелатонин, регулирующий пигментный обмен организма и оказывающий антигонадотропное действие. Кровоснабжение эпифиза осуществляется по кровеносной сети, образованной вторичными ветвями средней и задней мозговых артерий. Войдя в соединительнотканную капсулу органа, сосуды распадаются на множество капилляров органа с образованием сети, характеризующейся большим количеством анастомозов, Кровь от эпифиза отводится частично в систему большой мозговой вены Галена, некоторое количество ее поступает в вены сосудистого сплетения III желудочка. Нейросекреция эпифиза зависит от освещенности. Главным звеном в этой цепи является передний гипоталамус (супрахиазматическое ядро), которое получает прямой вход от волокон зрительного нерва. Далее от нейронов этого ядра образуется нисходящий путь к верхнему симпатическому узлу и затем в составе специального (пинеального) нерва поступает в эпифиз.
На свету продукция нейрогормонов в эпифизе угнетается, тогда как в течение темной фазы суток она усиливается. Мелатонин влияет на функции многих отделов центральной нервной системы и некоторые поведенческие реакции. Например, у человека инъекция мелатонина вызывает сон.
Другим физиологически активным веществом эпифиза, претендующим на роль нейрогормона, является серотонин — предшественник мелатонина. Исследования на животных показали, что содержание серотонина в эпифизе выше, чем в других органах, и зависит от вида, возраста животных, а также светового режима; оно подвержено суточным колебаниям с максимальным уровнем в дневное время. Суточная ритмика содержания серотонина в эпифизе возможна только в условиях целостности симпатической иннервации органа. В эпифизе содержится также значительное количество дофамина, который рассматривается в настоящее время как возможное физиологически активное вещество эпифиза. Перейдем к рассмотрению регуляций некоторых основных биологических мотиваций.

