 2015-04-17
2015-04-17 7610
7610Рецепторы делят на внешние, или экстероцепторы, и внутренние, или интерорецепторы. Экстероцепторы расположены на внешней поверхности тела животного или человека и воспринимают раздражения из внешнего мира (световые, звуковые, термические и др.). Интероцепторы находятся в различных тканях и внутренних органах (сердце, лимфатические и кровеносные сосуды, лёгкие и т.д.); воспринимают раздражители, сигнализирующие о состоянии внутренних органов (висцероцепторы), а также о положении тела или его частей в пространстве (вестибулоцепторы). Разновидность интероцепторов -- проприорецепторы, расположенные в мышцах, сухожилиях и связках и воспринимающие статическое состояние мышц и их динамику. В зависимости от природы воспринимаемого адекватного раздражителя различают механорецепторы, фоторецепторы, хеморецепторы, терморецепторы и др. У дельфинов, летучих мышей и ночных бабочек обнаружены рецепторы, чувствительные к ультразвуку, у некоторых рыб -- к электрическим полям. Менее изучен вопрос о существовании у некоторых птиц и рыб рецепторов, чувствительных к магнитным полям. Мономодальные рецепторы воспринимают раздражения только одного рода (механическое, световое или химическое); среди них -- рецепторы, различные по уровню чувствительности и отношению к раздражающему стимулу. Так, фоторецепторы позвоночных подразделяются на более чувствительные палочковые клетки, функционирующие как рецепторы сумеречного зрения, и менее чувствительные колбочковые клетки, обеспечивающие у человека и ряда животных дневное светоощущение и цветовое зрение; механорецепторы кожи -- на более чувствительные фазные рецепторы реагирующие только на динамическую фазу деформации, и статические, реагирующие и на постоянную деформацию, и т.д. В результате такой специализации рецепторы выделяются наиболее значительные свойства стимула и осуществляется тонкий анализ воспринимаемых раздражений. Полимодальные рецепторы реагируют на раздражения разного качества, например химическое и механическое, механическое и температурное. При этом закодированная в молекулах специфическая информация передаётся в центральную нервную систему по одним и тем же нервным волокнам в виде нервных импульсов, подвергаясь на своём пути неоднократному энергетическому усилению. Исторически сохранилось деление рецепторов на дистантные (зрительные, слуховые, обонятельные), воспринимающие сигналы от источника раздражения, находящегося на некотором расстоянии от организма, и контактные -- при непосредственном соприкосновении с источником раздражения. Различают также рецепторы первичные (первичночувствующие) и вторичные (вторичночувствующие). У первичных рецепторов субстрат, воспринимающий внешнее воздействие, заложен в самом сенсорном нейроне, который непосредственно (первично) возбуждается раздражителем. У вторичных рецепторах между действующим агентом и сенсорным нейроном располагаются дополнительные, специализированные (рецептирующие) клетки, в которых преобразуется (трансформируется) в нервные импульсы энергия внешних раздражений.
Все рецепторы характеризуются рядом общих свойств. Они специализированы для рецепции определённых, свойственных им раздражений, называемыми адекватными. При действии раздражений в рецепторах возникает изменение разности биоэлектрических потенциалов на клеточной мембране, так называемый рецепторный потенциал, который либо непосредственно генерирует ритмические импульсы в рецепторной клетке, либо приводит к их возникновению в другом нейроне, связанном с рецептором посредством синапса. Частота импульсов возрастает с увеличением интенсивности раздражения. При продолжительном действии раздражителя снижается частота импульсов в волокне, отходящем от рецептора; подобное явление уменьшения активности рецептора называется адаптацией физиологической. Для различных рецепторов время такой адаптации неодинаково. Рецепторы отличаются высокой чувствительностью к адекватным раздражителям, которая измеряется величиной абсолютного порога, или минимальной интенсивностью раздражения, способного привести рецепторы в состояние возбуждения. Так, например, 5--7 квантов света, падающего на рецептор глаза, вызывают световое ощущение, а для возбуждения отдельного фоторецептора достаточно 1 кванта. Рецептор можно возбудить и неадекватным раздражителем. Воздействуя, например, на глаз или ухо электрическим током, можно вызвать ощущение света или звука. Ощущения связаны со специфической чувствительностью рецептора, возникшей в ходе эволюции органической природы. Образное восприятие мира связано преимущественно с информацией, идущей с экстероцепторов. Информация с интероцепторов не приводит к возникновению чётких ощущений. Функции различных рецепторов взаимосвязаны. Взаимодействие вестибулярных рецепторов, а также рецепторов кожи и проприоцепторов со зрительными осуществляется центральной нервной системой и лежит в основе восприятия величины и формы предметов, их положения в пространстве. Рецепторы могут взаимодействовать между собой и без участия центральной нервной системы, т. е. вследствие непосредственной связи друг с другом. Такое взаимодействие, установленное на зрительных, тактильных и других рецепторов, имеет важное значение для механизма пространственно-временного контраста. Деятельность рецепторов регулируется центральной нервной системой, осуществляющей их настройку в зависимости от потребностей организма. Эти влияния, механизм которых изучен недостаточно, осуществляются посредством специальных эфферентных волокон, подходящих к некоторым рецепторным структурам.
Функции рецепторов исследуют методом регистрации биоэлектрических потенциалов непосредственно от рецепторов или связанных с ним нервных волокон, а также методом регистрации рефлекторных реакций, возникающих при раздражении рецепторов.
Рецепторы фармакологические (РФ), рецепторы клеточные, рецепторы тканевые, расположены на мембране эффекторной клетки; воспринимают регуляторные и пусковые сигналы нервной и эндокринной систем, действие многих фармакологических препаратов, избирательно влияющих на эту клетку, и трансформируют указанные воздействия в её специфическую биохимическую или физиологическую реакцию. Наиболее исследованы РФ, посредством которых осуществляется действие нервной системы. Влияние парасимпатического и двигательного отделов нервной системы (медиатор ацетилхолин) передают два типа РФ: Н-холиноцепторы передают нервные импульсы на скелетные мышцы и в нервных ганглиях с нейрона на нейрон; М-холино-цепторы участвуют в регуляции работы сердца и тонуса гладких мышц. Влияние симпатической нервной системы (медиатор норадреналин) и гормона мозгового вещества надпочечника (адреналина) передаётся альфа- и бета-адреноцепторами. Возбуждение альфа-адреноцепторов вызывает сужение сосудов, подъём артериального давления, расширение зрачка, сокращение ряда гладких мышц и т.д.; возбуждение бета-адреноцепторов -- увеличение сахара в крови, активацию ферментов, расширение сосудов, расслабление гладких мышц, усиление частоты и силы сердечных сокращений и т.д. Т. о., функциональное влияние осуществляется через оба типа адреноцепторов, а метаболическое -- преимущественно через бета-адреноцепторы. Обнаружены также РФ, чувствительные к дофамину, серотонину, гистамину, полипептидам и другим эндогенным биологически активным веществам и к фармакологическим антагонистам некоторых из этих веществ. Терапевтический эффект ряда фармакологических препаратов обусловлен их специфическим действием на специфические рецепторы.
15. Преобразование энергии раздражителя в рецепторах. Рецепторный и генераторный потенциалы. Закон Вебера-Фехнера. Абсолютный и дифференциальный пороги чувствительности.
В результате действия адекватного раздражителя у большинства рецепторов увеличивается проницаемость клеточной мембраны для катионов, что приводит к ее деполяризации. Исключением из общего правила являются фоторецепторы, где после поглощения энергии квантов света в связи особенностями управления ионными каналами происходит гиперполяризация мембраны. Изменение величины мембранного потенциала рецепторов в ответ на действие стимула представляет собой рецепторный потенциал — входной сигнал первичных сенсорных нейронов. Если величина рецепторного потенциала достигнет критического уровня деполяризации или превысит его, генерируются потенциалы действия, с помощью которых сенсорные нейроны передают в центральную нервную систему информацию о действующих стимулах.
Генерация потенциалов действия происходит в ближайшем к рецепторам перехвате Ранвье миелинизированных волокон или ближайшей к рецепторам части мембраны безмиелинового волокна. Минимальная сила адекватного стимула, достаточная для генерации потенциалов действия в первичном сенсорном нейроне, определяется как его абсолютный порог. Минимальный прирост силы стимула, сопровождающийся значимым изменением реакции сенсорного нейрона, представляет собой дифференциальный порог его чувствительности.
Вебер (1831) и Фехнер (1860) доказали зависимость между абсолютным порогом и дифференциальным порогом стимула.
JD/J=K
J- исходный раздражитель
JD- прирост раздражения
K- постоянная величина
Информация о силе действующего на рецепторы стимула кодируется двумя способами: частотой потенциалов действия, возникающих в сенсорном нейроне (частотное кодирование), и числом сенсорных нейронов, возбудившихся в ответ на действие стимула. При увеличении силы действующего на рецепторы раздражителя повышается амплитуда рецепторного потенциала, что, как правило, сопровождается увеличением частоты потенциалов действия в сенсорном нейроне первого порядка. Чем шире имеющийся частотный диапазон потенциалов действия у сенсорных нейронов, тем большее число промежуточных значений силы раздражителя способна различать сенсорная система. Первичные сенсорные нейроны одинаковой модальности различаются порогом возбуждения, поэтому при действии слабых стимулов возбуждаются только наиболее чувствительные нейроны, но с увеличением силы раздражителя на него реагируют и менее чувствительные нейроны, имеющие более высокий порог раздражения. Чем больше первичных сенсорных нейронов возбудится одновременно, тем сильнее будет их совместное действие на общий нейрон второго порядка, что в итоге отразится на субъективной оценке интенсивности действующего раздражителя.
Длительность ощущения зависит от реального времени между началом и прекращением воздействия на рецепторы, а также от их способности уменьшать или даже прекращать генерацию нервных импульсов при продолжительном действии адекватного стимула. При длительном действии стимула порог чувствительности рецепторов к нему может повышаться, что определяется как адаптация рецепторов. Механизмы адаптации не одинаковы в рецепторах разных модальностей, среди них различают быстро адаптирующиеся (например, тактильные рецепторы кожи) и медленно адаптирующиеся рецепторы (например, проприоцепторы мышц и сухожилий). Быстро адаптирующиеся рецепторы сильнее возбуждаются в ответ на быстрое нарастание интенсивности стимула (фазический ответ), а их быстрая адаптация способствует освобождению восприятия от биологически незначительной информации (например, контакт между кожей и одеждой). Возбуждение медленно адаптирующихся рецепторов мало зависит от скорости изменения стимула и сохраняется при его длительном действии (тонический ответ), поэтому, например, медленная адаптация проприоцепторов позволяет человеку получать нужную ему для сохранения позы информацию в течение всего необходимого времени.
Существуют сенсорные нейроны, генерирующие потенциалы действия спонтанно, т. е. при отсутствии раздражения (например, сенсорные нейроны вестибулярной системы), такая активность называется фоновой. Частота нервных импульсов в этих нейронах может увеличиваться или уменьшаться в зависимости от интенсивности действующего на вторичные рецепторы стимула, кроме того, она может определяться направлением, в котором отклоняются чувствительные волоски механорецепторов. Например, отклонение волосков вторичных механорецепторов в одну сторону сопровождается повышением фоновой активности сенсорного нейрона, которому они принадлежат, а в противоположную сторону — понижением его фоновой активности. Указанный способ рецепции позволяет получать информацию и об интенсивности стимула, и о направлении, в котором он действует.
16. Кодирование информации в сенсорных системах.
Кодирование - процесс преобразования информации в условную форму (код), удобную для передачи по каналу связи. Универсальным кодом нервной системы являются нервные импульсы, которые распространяются по нервным волокнам. При этом содержание информации определяется не амплитудой импульсов (они подчиняются закону «Все или ничего»), а частотой импульсов (интервалами времени между отдельными импульсами), объединением их в пачки, числом импульсов в пачке, интервалами между пачками. Передача сигнала от одной клетки к другой во всех отделах анализатора осуществляется с помощью химического кода, т.е. различных медиаторов. Для хранения информации в ЦНС кодирование осуществляется с помощью структурных изменений в нейронах (механизмы памяти). Кодируемые характеристики раздражителя. В анализаторах кодируются качественная характеристика раздражителя (например, свет, звук), сила раздражителя, время его действия, а также пространство, т.е. место действия раздражителя и локализация его в окружающей среде. В кодировании всех характеристик раздражителя принимают участие все отделы анализатора.
В периферическом отделе анализатора кодирование качества раздражителя (вид) осуществляется за счет специфичности рецепторов, т.е. способности воспринимать раздражитель определенного вида, к которому он приспособлен в процессе эволюции, т.е. к адекватному раздражителю. Так, световой луч возбуждает только рецепторы сетчатки, другие рецепторы (обоняния, вкуса, тактильные и т.д.) на него обычно не реагируют.
Сила раздражителя может кодироваться изменением частоты импульсов в генерируемых рецепторами при изменении силы раздражителя, что определяется общим количеством импульсов в единицу времени. Это так называемое частотное кодирование.
Пространство кодируется величиной площади, на которой возбуждаются рецепторы, это пространственное кодирование. Время действия раздражителя на рецептор кодируется тем, что он начинает возбуждаться с началом действия раздражителя и прекращает возбуждаться сразу после выключения раздражителя (временное кодирование).
В проводниковом отделе анализатора кодирование осуществляется только на «станциях переключения», т. е. при передаче сигнала от одного нейрона к другому, где происходит смена кода. В нервных волокнах информация не кодируется, они исполняют роль проводов, по которым передается информация, закодированная в рецепторах и переработанная в центрах нервной системы. Между импульсами в отдельном нервном волокне могут быть различные интервалы, импульсы формируются в пачки с различным числом, между отдельными пачками могут быть также различные интервалы. Все это отражает характер закодированной в рецепторах информации. В нервном стволе при этом может изменяться также число возбужденных нервных волокон, что определяется изменением числа возбужденных рецепторов или нейронов на предыдущем переходе сигнала с одного нейрона на другой. На станциях переключения, например в зрительном бугре, информация кодируется, во-первых, за счет изменения объема импульсации на входе и на выходе, а во-вторых, за счет пространственного кодирования, т.е. за счет связи определенных нейронов с определенными рецепторами. В обоих случаях
чем сильнее раздражитель, тем большее число нейронов возбуждается.
В корковом конце анализатора происходит частотно-пространственное кодирование, нейрофизиологической основой которого является пространственное распределение ансамблей специализированных нейронов и их связей с определенными видами рецепторов. Импульсы поступают от рецепторов в определенные зоны коры с различными временными интервалами. Поступающая в виде нервных импульсов информация перекодируется в структурные и биохимические изменения в нейронах (механизмы памяти). В коре мозга осуществляется высший анализ и синтез поступившей информации. Анализ заключается в том, что с помощью возникающих ощущений мы различаем действующие раздражители (качественно – свет, звук и т.д.) и определяем силу, время и место, т.е. пространство, на которое действует раздражитель, а также его локализацию (источник звука, света, запаха). Синтез реализуется в узнавании известного предмета, явления или в формировании образа, впервые встречаемого предмета, явления.
Итак, процесс передачи сенсорного сообщения сопровождается многократным перекодированием и завершается высшим анализом и синтезом, который происходит в корковом отделе анализаторов. После этого уже происходит выбор или разработка программы ответной реакции организма.
17. Структурно-функциональная характеристика коры больших полушарий. Локализация функций в коре больших полушарий.
18. Рецептивное поле. Топическая организация сенсорных систем.
Рецептивное поле (англ. receptive field) сенсорного нейрона — участок с рецепторами, которые при воздействии на них определённого стимула приводят к изменению возбуждения этого нейрона.
Концепция рецептивных полей может быть применима ко всей нервной системе. Если множество сенсорных рецепторов образуют синапсы c единственным нейроном, они совместно формируют рецептивное поле этого нейрона. Например, рецептивное поле ганглионарной (ганглиозной) клетки сетчатки глаза представлено фоторецепторными клетками (англ.)русск. (палочками или колбочками), а группа ганглионарных клеток в свою очередь создаёт рецептивное поле для одного из нейронов мозга. В итоге к одному нейрону более высокого синаптического уровня сходятся импульсы от многих фоторецепторов; и этот процесс называется конвергенцией. Рецептивным полем называется область, занимаемая совокупностью всех рецепторов, стимуляция которых приводит к возбуждению сенсорного нейрона (рис. 17.1). Максимальная величина рецептивного поля первичного сенсорного нейрона определяется пространством, которое занимают все ветви его периферического отростка, а число рецепторов, имеющихся в этом пространстве, указывает на плотность иннервации. Высокая плотность иннервации сочетается, как правило, с малыми размерами рецептивных полей и, соответственно, высоким пространственным разрешением, позволяющим различать стимулы, действующие на соседние рецептивные поля. Малые рецептивные поля типичны, например, для центральной ямки сетчатки и для пальцев рук, где плотность рецепторов значительно выше, чем на периферии сетчатки или в коже спины, для которых характерны большая величина рецептивных полей и меньшее пространственное разрешение. Рецептивные поля соседних сенсорных нейронов могут частично перекрывать друг друга, поэтому информация о действующих на них стимулах передается не по одному, а по нескольким параллельным аксонам, что повышает надежность ее передачи.
Рис. 17.1. Рецептивные поля первичных сенсорных нейронов и сенсорных нейронов второго порядка.
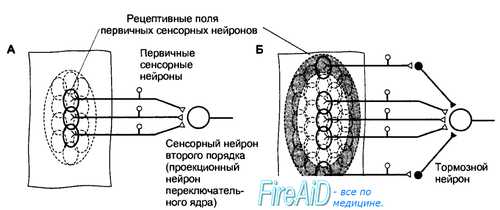
А. Рецептивные поля первичных сенсорных нейронов ограничены областью их чувствительных окончаний. Рецептивное поле переключательного нейрона образуется из суммы рецептивных полей конвергирующих к нему первичных сенсорных нейронов.
Б. Раздражение центральной или периферической области рецептивного поля сенсорного нейрона второго и следующих порядков сопровождается противоположным эффектом. Как видно на схеме, раздражение центра рецептивного поля вызовет возбуждение проекционного нейрона, а раздражение периферической области — торможение с помощью интернейронов переключательного ядра (латеральное торможение). В результате контраста, создаваемого между центром и периферией рецептивного поля, выделяется информация для передачи на следующий иерархический уровень.
Величина рецептивных полей сенсорных нейронов второго и следующих порядков больше, чем у первичных сенсорных нейронов, поскольку центральные нейроны получают информацию от нескольких конвергирующих к ним нейронов предшествующего уровня. От центра рецептивного поля информация передается непосредственно к сенсорным нейронам следующего порядка, а от периферии — к тормозным интернейронам переключательного ядра, поэтому центр и периферия рецептивного поля являются реципрокными по отношению друг к другу. В результате сигналы от центра рецептивного поля беспрепятственно достигают следующего иерархического уровня сенсорной системы, тогда как сигналы, поступающие от периферии рецептивного поля, тормозятся (в другом варианте организации рецептивного поля легче пропускаются сигналы от периферии, а не от центра). Такая функциональная организация рецептивных полей обеспечивает выделение наиболее значимых сигналов, легко различаемых на контрастном с ними фоне.
Сенсорный проводящий путь состоит из ряда модально- специфических нейронов, которые соединены синапсами. Такой принцип организации получил название меченой линии или топической организации. Суть этого принципа заключается в пространственно упорядоченном расположении нейронов на различных уровнях сенсорных систем соответственно характеристикам их рецептивных полей.
Рецептивное поле с морфологической точки зрения — это участок рецепторной поверхности, с которым данная нервная структура (волокно, нейрон) связана анатомически (жестко). С функциональной точки зрения — рецептивное поле — понятие динамическое, означающее, что один и тот же нейрон в различные отрезки времени в зависимости, например, от характеристики воздействия может оказаться связанным с различным числом рецепторов.
Принципу меченой линии противопоставлялась теория “ структуры ответа”, согласно которой рецепторы кодируют качественные особенности раздражителей структурой импульсного ответа. Эта теория предполагала отсутствие жестких связей между рецепторами и центральными нейронами. Основанием для нее послужили экспериментальные данные, показавшие, что кодирование информации осуществляется не одиночными импульсами, а группой равномерно следующих потенциалов действия. В качестве сигнальных признаков могут быть использованы дополнительные параметры активности рецепторов, например, частота импульсации или продолжительность межимпульсных интервалов.
Для равномерно следующих импульсов сигнальными признаками могут служить число импульсов в пачке или продолжительность пачек, а также интервалы между ними и периодичность их следования. Такое кодирование открывает безграничные возможности, т. к. вероятны самые разнообразные вариации с пачками импульсов. Пространственно- временное распределение электрической активности нервных волокон называют паттернами. Разнообразные качества стимулов, согласно этой теории, отображаются характерными “ узорами” паттернов. Нейроны способны расшифровать эти сигналы и в зависимости от их структуры формировать ощущение, которое соответствует раздражителю, кодируемого определенными паттернами.
Нейрон, по- разному реагируя на различные паттерны, может участвовать в выполнении нескольких функций. Каждый оттенок качества ощущения возникает в результате деятельности комплекса нейронов, образующих динамические ансамбли, формирование которых зависит от характера паттернов, приходящих от рецепторов.
Для каждой модальности имеется своя форма кодирования информации в соответствии с физическими свойствами различаемых стимулов. Одни качества распознаются сенсорными системами, функционирующими по принципу топической организации, другие кодируются паттернами. Например, распознавание многих качеств зрительных образов осуществляется меченными линиями, а вкусовые раздражители кодируются паттернами.
19. Рефлекторная дуга.
Структурную основу рефлекторной деятельности составляют нейронные цепи из рецепторных, вставочных и эффекторных нейронов. Они образуют путь, по которому проходят нервные импульсы от рецептора к исполнительному органу при осуществлении всякого рефлекса. Этот путь носит название рефлекторной дуги. В ее состав входят:
1. воспринимающие раздражения рецепторы;
2. афферентные нервные волокна - отростки рецепторных нейронов, несущие возбуждение к центральной нервной системе;
3. нейроны и синапсы, передающие импульсы к эффекторным нейронам;
4. эфферентные нервные волокна, проводящие импульсы от центральной нервной системы на периферию;
5. исполнительный орган, деятельность которого изменяется в результате рефлекса.
Простейшего рефлекторную дугу можно схематически представить себе как образованную всего двумя нейронами: рецепторным и эффекторным, между которыми имеется один синапс. Такую рефлекторную дугу называют двунейронной и моносинаптической.
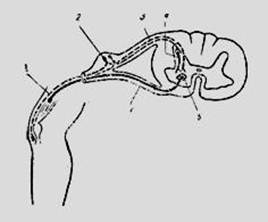
Рефлекторные дуги большинства рефлексов включают не два, а большее число нейронов: рецепторный, один или несколько вставочных и эффекторный. Такие рефлекторные дуги называют многонейронными и полисинаптическими. Возможны различные варианты полисинаптических рефлекторных дуг. Такая наиболее простая дуга включает в свой состав всего три нейрона и два синапса между ними. Существуют полисинаптические рефлекторные дуги, в которых рецепторный нейрон соединен с несколькими вставочными, каждый из которых образует синапсы на разных или на одном и том же эффекторном нейроне.
В периферической нервной системе различают рефлекторные дуги (нейронные цепи)
· соматической нервной системы, иннервирующие скелетную иускулатуру
· вегетативной нервной системы, иннервирующие внутренние органы: сердце, желудок, кишечник, почки, печень и т.д.
Рефлекторная дуга состоит из пяти отделов:
1. рецепторов, воспринимающих раздражение и отвечающих на него возбуждением. Рецепторами могут быть окончания длинных отростков центростремительных нервов или различной формы микроскопические тельца из эпителиальных клеток, на которых оканчиваются отростки нейронов. Рецепторы расположены в коже, во всех внутренних органах, скопления рецепторов образуют органы чувств (глаз, ухо и т. д.).
2. чувствительного (центростремительного, афферентного) нервного волокна, передающего возбуждение к центру; нейрон, имеющий данное волокно, также называется чувствительным. Тела чувствительных нейронов находятся за пределами центральной нервной системы - в нервных узлах вдоль спинного мозга и возле головного мозга.
3. нервного центра, где происходит переключение возбуждения с чувствительных нейронов на двигательные; Центры большинства двигательных рефлексов находятся в спинном мозге. В головном мозге расположены центры сложных рефлексов, таких, как защитный, пищевой, ориентировочный и т. д. В нервном центре происходит синаптическое соединение чувствительного и двигательного нейрона.
4. двигательного (центробежного, эфферентного) нервного волокна, несущего возбуждение от центральной нервной системы к рабочему органу; Центробежное волокно - длинный отросток двигательного нейрона. Двигательным называется нейрон, отросток которого подходит к рабочему органу и передает ему сигнал из центра.
5. эффектора - рабочего органа, который осуществляет эффект, реакцию в ответ на раздражение рецептора. Эффекторами могут быть мышцы, сокращающиеся при поступлении к ним возбуждения из центра, клетки железы, которые выделяют сок под влиянием нервного возбуждения, или другие органы.
Простейшую рефлекторную дугу можно схематически представить как образованную всего двумя нейронами: рецепторным и эффекторным, между которыми имеется один синапс. Такую рефлекторную дугу называют двунейронной и моносинаптической. Моносинаптические рефлекторные дуги встречаются весьма редко. Примером их может служить дуга миотатического рефлекса.
В большинстве случаев рефлекторные дуги включают не два, а большее число нейронов: рецепторный, один или несколько вставочных и эффекторный. Такие рефлекторные дуги называют многонейронными и полисинаптическими. Примером полисинаптической рефлекторной дуги является рефлекс отдергивания конечности в ответ на болевое раздражение.
Рефлекторная дуга соматической нервной системы на пути от ЦНС к скелетной мышце нигде не прерывается в отличии от рефлекторной дуги вегетативной нервной системы, которая на пути от ЦНС к иннервируемому органу обязательно прерывается с образованием синапса - вегетативного ганглия.
Вегетативные ганглии, в зависимости от локализации, могут быть разделены на три группы:
1. позвоночные (вертебральные) ганглии - относятся к симпатической нервной системе. Они расположены по обе стороны позвоночника, образуя два пограничных ствола (их еще называют симпатическими цепочками)
2. предпозвоночные (превертебральные) ганглии располагаются на большем расстояни от позвоночника, вместе с тем они находятся в некотором отдалении и от иннервируемых ими органов. К числу превертебральных ганглиев относят ресничный узел, верхний и средний шейный симпатические узлы, солнечное сплетение, верхний и нижний брыжеечные узлы.
3. внутриорганные ганглии расположены во внутренних органах: в мышечных стенках сердца, бронхов, средней и нижней трети пищевода, желудка, кишечника, желчного пузыря, мочевого пузыря, а также в железах внешней и внутренней секреции. На клетках этих ганглий прерываются парасимпатические волокна.
Такое различие соматической и вегетативной рефлекторной дуги обусловлено анатомическим строением нервных волокон, составляющих нейронную цепь, и скоростью проведения по ним нервного импульса.
Для осуществления любого рефлекса необходима целостность всех звеньев рефлекторной дуги. Нарушение хотя бы одного из них ведет к исчезновению рефлекса.
20. Безусловные рефлексы, их характеристика. Инстинкты.
Безусловные рефлексы — это врожденная видоспецифическая реакция организма, рефлекторно возникающая в ответ на специфическое воздействие раздражителя, на воздействие биологически значащего (боль, пища, тактильное раздражение и т. д.) стимула, адекватного для данного вида деятельности.
Безусловные рефлексы:
· Врожденные наследственно передающиеся реакции, большинство из них начинают функционировать сразу после рождения.
· Являются видовыми, т.е. свойственны всем представителям данного вида.
· Постоянны и сохраняются в течение всей жизни.
· Осуществляются за счет низших отделов ЦНС (подкорковые ядра, ствол мозга, спинной мозг).
· Возникают в ответ на адекватные раздражения, действующие на определенное рецептивное поле.
По уровню сложности безусловные рефлексы подразделяются на:
· простые безусловные рефлексы
· рефлекторные акты
· реакции поведения
· инстинкты
Простые безусловные рефлексы - элементарные врожденные реакции на раздражители. Например, отдергивание конечности от горячего предмета, мигание века при попадании в глаз соринки и т.д. Простые безусловные рефлексы на соответствующий раздражитель проявляются всегда, изменению и исправлению не поддаются.
Рефлекторные акты - определяемые несколькими простыми безусловными рефлексами действия, выполняемые всегда одинаково и независимо от сознания собаки. В основном рефлекторные акты обеспечивают жизнедеятельность организма, поэтому всегда проявляются надежно и не поддаются коррекции.
Некоторые примеры рефлекторных актов:
- дыхание;
- глотание;
- отрыгивание
Дрессируя и воспитывая собаку, следует помнить, что единственный способ предотвратить проявление того или иного рефлекторного акта - изменить или убрать раздражитель, его вызывающий. Так, если вы хотите, чтобы во время отработки навыков послушания ваш питомец не отправлял естественные надобности (а он все равно это сделает, если есть необходимость, невзирая на ваш запрет, ведь это проявление рефлекторного акта), то выгуляйте собаку перед тренировкой. Тем самым вы устраните соответствующие раздражители, вызывающие рефлекторный акт, нежелательный для вас.
Реакции поведения - стремление собаки осуществить те или иные действия, основанное на комплексе рефлекторных актов и простых безусловных рефлексов.
Таким образом, реакции поведения являются причиной многих действий собаки, но в реальной обстановке их проявление можно контролировать. Мы привели негативный пример, показывающий нежелательное поведение собаки. Но попытки выработать нужное поведение при отсутствии необходимых реакций окончатся неудачей. Например, бесполезно готовить розыскную собаку из кандидата, у которого отсутствует обонятельно-поисковая реакция. Не получится охранника из собаки с пассивно-оборонительной реакцией (из трусливой собаки).
Инстинкт – это врожденная, строго постоянная, специфическая для каждого вида форма приспособительного поведения, побуждаемая основными биологическими потребностями организма и специфическими раздражителями внешней среды.
Естественный отбор влияет на поведение таким же образом, как и на строение тела, его окраску и на все остальные морфологические и физиологические признаки и свойства организмов.
При относительно небольшой сложности мозга естественный отбор ведет к совершенствования жестко запрограммированных форм поведения, обеспечивающих выживание.
В целом, естественный отбор вел к появлению организмов со все более сложным и гибким поведением, обеспечивающим выживание в изменчивых условиях. Как следствие этой тенденции на земле появился человек.
Критерии и признаки инстинктов:
1) Побуждение (мотивация) и способность к действию принадлежит к числу наследственных свойств вида;
2) такие действия не требуют предварительного обучения (хотя обучение может развивать и совершенствовать его выполнение!);
3) выполняются по существу одинаково у всех нормальных представителей вида;
4) связаны с нормальным функционированием его органов (например, инстинкт рытья нор сочетается с соответствующим строением лап, приспособленных для рытья);
5) приспособлены к экологическим условиям обитания вида (т.е. обеспечивают выживание в конкретных экологических условиях).
Уровни рефлекторных поведенческих реакций (по А.Б. Когану)
· Первый уровень: элементарные безусловные рефлексы. Это простые безусловно-рефлекторные реакции, осуществляются на уровне отдельных сегментов спинного мозга. Реализуются в соответствии с генетически детерминированными программами. Стереотипны. Осуществляются неосознанно.
· Второй уровень: координационные безусловные рефлексы. Это сложные акты сокращения и расслабления различных мышц или возбуждения и торможения функций внутренних органов, причем эти реципрокные отношения хорошо координированы.
В осуществлении координации безусловных рефлексов большое значение имеют обратные связи.
Формируются на базе элементарных безусловных рефлексов (первого уровня рефлекторных реакций).
Это локомоторные акты и вегетативные процессы, направленные на поддержание гомеостаза.
· Третий уровень организации рефлекторных реакций –интегративные безусловные рефлексы.
Возникают под действием биологически важных стимулов (пищевых и болевых).
Интегративные безусловные рефлексы – комплексные поведенческие акты, имеют системный характер с выраженными соматическим и вегетативными компонентами. Например, локомоторные акты сопровождаются усилением кровообращения, дыхания и т.д.
· Четвертый уровень – сложнейшие безусловные рефлексы (инстинкты).
Впервые предположил, что инстинкты – тоже рефлексы, Герберт Спенсер.
Сложнейшие безусловные рефлексы выполняются по генетически заданным программам, пусковой раздражитель запускает их целиком.
· Пятый уровень – элементарные условные рефлексы.
Они вырабатываются в процессе индивидуальной жизни.
В раннем возрасте формируются простые условно-рефлекторные реакции. С течением жизни они усложняются. В образовании условных рефлексов участвует кора больших полушарий.
Условно-рефлекторный механизм поведения отличается высокой степенью надежности, которая обеспечивается многоканальностью и взаимозаменяемостью нервных связей в пластичных структурах ЦНС
· Шестой уровень поведенческих актов - сложные формы психической деятельности.
В основе – интеграция элементарных условных рефлексов и аналитико-синтетических механизмов абстрагирования.
21. Условные рефлексы, их характеристика.
Условный рефлекс – это сложная многокомпонентная реакция, которая вырабатывается на базе безусловных рефлексов с использованием предшествующего индифферентного раздражителя. Он имеет сигнальный характер и организм встречает воздействие безусловного раздражителя подготовленным.
Условные рефлексы:
· Реакции, приобретенные в процессе индивидуальной жизни.
· Индивидуальны.
· Непостоянны – могут возникать и исчезать.
· Являются преимущественно функцией коры больших полушарий.
· Возникают на любые раздражители, действующие на разные рецептивные поля.
Классификация условных рефлексов
· По степени близости сигнального стимула к биологии животного:
· Натуральные условные рефлексы
· Искусственные условные рефлексы
На основе локализации и свойствах афферентного звена дуги условного рефлекса:
· Экстероцептивные
· Интероцептивные
· Проприоцептивные
По модальности адекватного раздражителя:
Механо-, фото-, хемо-, термо-, осморецепторные условные рефлексы.
· Свойства эфферентного звена условных рефлексов:
· Вегетативные
· Соматические
По степени (глубине) абстрагирования:
· Условные рефлексы I, II и более высоких порядков.
· Третичный условный рефлекс в 20-х годах 20 века выработал сотрудник И.П.Павлова – Д.С.Фурсиков.
· Рефлекс IV порядка нельзя выработать у собак, но можно у дельфинов.
· У лошадей глубина абстрагирования – рефлексы V – VI порядков.
По структуре:
· простые и сложные
По соотношению во времени сигнала и подкрепления:
· Наличные (подкрепляющий стимул подается во время действия сигнального раздражителя).
· Следовые (делают паузу между окончанием действия условного раздражителя и началом подкрепления; по мере усложнения эксперимента пауза – от 15-20с до 4-5 минут).
Условные рефлексы – это рефлексы на будущие события. Биологическое значение условных
рефлексов состоит в их предупредительной роли, они имеют для организма приспособительное
значение, готовя организм к будущей полезной поведенческой деятельности и помогая ему избежать вредных воздействий, тонко и эффективно адаптироваться к окружающей природной и социальной среде. Условные рефлексы формируются благодаря пластичности нервной системы.
22. Правила выработки условных рефлексов.
Для выработки условного рефлекса необходимо:
1) наличие двух раздражителей, один из которых безусловный (пища, болевой раздражитель и др.), вызывающий безусловно-рефлекторную реакцию, а другой - условный (сигнальный), сигнализирующий о предстоящем безусловном раздражении (свет, звук, вид пищи и т.д.);
2) многократное сочетание условного и безусловного раздра жителей (хотя возможно образование условного рефлекса при их однократном сочетании);
3) условный раздражитель должен предшествовать действию безусловного;
4) в качестве условного раздражителя может быть использован любой раздражитель внешней или внутренней среды, который должен быть по возможности индифферентным, не вызывать обронительной реакции,не обладать чрезмерной силой и способен привлекать внимание;
5) безусловный раздражитель должен быть достаточно сильным, в противном случае временная связь не сформируется;
6) возбуждение от безусловного раздражителя должно быть более сильным, чем от условного;
7) необходимо устранить посторонние раздражители, так как они могут вызывать торможение условного рефлекса;
8) животное, у которого вырабатывается условный рефлекс, должно быть здоровым;
9) при выработке условного рефлекса должна быть выражена мотивация, например, при выработке пищевого слюноотделительного рефлекса животное должно быть голодным, у сытого - этот рефлекс не вырабатывается.
Условные рефлексы легче вырабатывать на экологически близкие данному животному воздействия. В связи с этим условные рефлексы делятся на натуральные и искусственные. Натуральные условные рефлексы вырабатываются на агенты, которые в естественных условиях действуют вместе с раздражителем, вызывающим безусловный рефлекс (например, вид пищи, ее запах и т.д.). Все остальные условные рефлексы искусственные, т.е. вырабатываются на агенты, в норме не связанные с действием безусловного раздражителя, например, пищевой слюноотделительный рефлекс на звонок.
Физиологической основой для возникновения условных рефлексов служит образование функциональных временных связей в высших отделах ЦНС. Временная связь - это совокупность нейрофизиологических, биохимических и ультраструктурных изменений в мозге, возникающих в процессе совместного действия условного и безусловного раздражителей. И.П.Павлов высказал предположение, что при выработке условного рефлекса происходит формирование временной нервной связи между двумя группами клеток коры - корковыми представительствами условного и безусловного рефлексов. Возбуждение от центра условного рефлекса может передаваться к центру безусловного рефлекса от нейрона к нейрону.
Следовательно, первый путь образования временной связи между корковыми представительствами условного и безусловного рефлексов является внутрикорти-кальным. Однако при разрушении коркового представительства условного рефлекса выработанный условный рефлекс сохраняется. По-видимому, образование временной связи идет между подкорковым центром условного рефлекса и корковым центре-безусловного рефлекса. При разрушении коркового представительства безусловного рефлекса условный рефлекс также сохраняется. Следовательно, выработка временной связи может идти между корковым центром условного рефлекса и подкорковьг центром безусловного рефлекса.
Разобщение корковых центров условного и безусловного рефлексов путем пересечения коры мозга не препятствует образованию условного рефлекса. Это свидетельствует о том, что временная связь может образоватьс между корковым центром условного рефлекса, подкорковьг центром безусловного рефлекса и корковым центром безусловного рефлекса.
Имеются различные мнения по вопросу о механизмах обрс зования временной связи. Возможно, образование временно связи происходит по принципу доминанты. Очаг возбуждения с безусловного раздражителя всегда сильнее, чем от условного, так как безусловный раздражитель всегда биологически более значим для животного. Этот очаг возбуждения является доминантным, следовательно притягивает к себе возбуждение от очага условного раздражения. Если возбуждение прошло по каким-либо нервным цепям, то в следующий раз оно по этим путям пройдет значительно легче (явление "проторения пути"). В основе этого лежат: суммация возбуждений, длительное повышение возбудимости синаптических образований, увеличение количества медиатора в синапсах, увеличение образования новых синапсов. Все это создает структурные предпосылки к облегчению движения возбуждения по определенным нейронным цепям.
Другим представлением о механизме формирования временной связи является конвергентная теория. В ее основе лежит способность нейронов отвечать на раздражения разных модальностей. По представлениям П.К.Анохина, условный и безусловный раздражители вызывают распространенную активацию корковых нейронов благодаря включению ретикулярной формации. В результате восходящие сигналы (условного и безусловного раздражителей) перекрываются, т.е. происходит встреча этих возбуждений на одних и тех же корковых нейронах. В результате конвергенции возбуждений возникают и стабилизируются временные связи между корковыми представительствами условного и безусловного раздражителей.
23. Условные рефлексы второго и более высокого порядка. Динамический стереотип.

