2015-04-20
2015-04-20 977
977Общее направление эволюции ЦНС — увеличение числа вставочных нейронов. Из более чем ста миллиардов нейронов человека не менее 70% составляют именно вставочные нервные клетки.
Среди беспозвоночных наиболее примитивный тип нервной системы в виде диффузной нервной сети встречается у кишечнополостных. Их нервная сеть представляет собой скопление мультиполярных и биполярных нейронов, отростки которых могут перекрещиваться, прилегать друг к другу и лишены функциональной дифференциации на аксоны и дендриты.
Эпидермальные нервные сплетения, напоминающие нервные сети кишечнополостных, могут быть обнаружены и у более высоко организованных беспозвоночных (плоские и кольчатые черви), однако здесь они занимают подчиненное положение по отношению к ЦНС, которая выделяется как самостоятельный отдел.
Ганглионизация нервных элементов получает развитие у высших беспозвоночных, кольчатых червей, моллюсков и членистоногих. У большинства кольчатых червей брюшные стволы ганглионизированы таким образом, что в каждом сегменте тела формируется по одной паре ганглиев, соединенных коннективами с другой парой, расположенной в соседнем сегменте.
Эволюция нервной системы беспозвоночных идет не только по пути концентрации нервных элементов, но и в направлении усложнения структурных взаимоотношений в пределах ганглиев. Не случайно брюшную нервную цепочку сравнивают со спинным мозгом позвоночных животных. Как и в спинном мозгу, в ганглиях обнаруживается поверхностное расположение проводящих путей, дифференциация на моторную, чувствительную и ассоциативные области.
Прогрессивное развитие мозга у головоногих моллюсков и насекомых создает предпосылку для возникновения своеобразной иерархии командных систем управления поведением. Низший уровень интеграции в сегментарных ганглиях насекомых служит основой для автономной деятельности и координации элементарных двигательных актов. В то же время мозг представляет собой следующий, более высокий уровень интеграции, где могут осуществляться межанализаторный синтез и оценка биологической значимости информации. На основе этих процессов формируются нисходящие команды, обеспечивающие вариантность запуска нейронов сегментарных центров. Очевидно, взаимодействие двух уровней интеграции лежит в основе пластичности поведения высших беспозвоночных, включающего врожденные и приобретенные реакции.
Нервная система позвоночных закладывается в виде сплошной нервной трубки, которая дифференцируется на различные отделы и является также источником периферических симпатических, парасимпатических и метасимпатических нервных узлов. У наиболее древних хордовых (бесчерепных) головной мозг отсутствует, и нервная трубка представлена в малодифференцированном состоянии.
В ходе дальнейшей эволюции наблюдается перемещение некоторых функций и систем интеграции из спинного мозга в головной — процесс энцефализации, формируется головной мозг как надстройка над системами дистантной рецепции.
Передний мозг круглоротых длительное время считали чисто обонятельным. Однако исследования недавнего времени показали, что обонятельные входы в передний мозг не являются единственными, а дополняются сенсорными входами других модальностей. Очевидно, уже на ранних этапах передний мозг начинает участвовать в переработке информации и управлении поведением. Вместе с тем энцефализация как магистральное направление развития мозга не исключает эволюционных преобразований в спинном мозгу круглоротых. Нейроны кожной чувствительности выделяются из спинного мозга и концентрируются в спинномозговой ганглий. Наблюдается совершенствование проводниковой части спинного мозга. Проводящие волокна боковых столбов имеют контакты с мощной дендритной сетью мотонейронов. Формируются нисходящие связи головного мозга со спинным через мюллеровские волокна — гигантские аксоны клеток, лежащих в среднем и продолговатом мозгу.
Наиболее существенные в эволюционном плане изменения происходят в промежуточном мозгу амфибий. Здесь обособляется таламус (зрительный бугор), дифференцируются структурированные ядра (наружное коленчатое тело) и восходящие пути, связывающие зрительный бугор с корой (таламокортикальный путь).
В полушариях переднего мозга происходит дальнейшая дифференциация зачатков старой и древней коры. В старой коре (археокортексе) обнаруживаются звездчатые и пирамидные клетки. В промежутке между старой и древней корой появляется полоска плаща, которая является предтечей новой коры (неокортекса).
В целом развитие переднего мозга создает предпосылки для перехода от свойственной рыбам системы интеграции к системе, где ведущим отделом становится передний мозг, а таламус промежуточного мозга превращается в коллектор всех афферентных сигналов.
Развитие таламокортикальной системы связей у рептилий приводит к формированию новых проводящих путей, как бы подтягивающихся к молодым формациям мозга.
В боковых столбах спинного мозга рептилий обособляется восходящий спинно—таламический путь, который проводит к головному мозгу информацию о температурной и болевой чувствительности. Здесь же в боковых столбах формируется новый нисходящий путь — красно—ядерно—спинномозговой. Он связывает мотонейроны спинного мозга с красным ядром среднего мозга. Эта многозвенная система объединяет влияние переднего мозга, мозжечка, ретикулярной формации ствола, ядер вестибулярного комплекса и координирует двигательную активность. У рептилий, как истинно наземных животных, возрастает роль зрительной и акустической информации, возникает необходимость сопоставления этой информации с обонятельной и вкусовой. В соответствии с этими биологическими изменениями в стволовой части мозга рептилий происходит целый ряд структурных изменений. В продолговатом мозгу дифференцируются слуховые ядра, помимо улиткового ядра появляется угловое, связанное со средним мозгом. В среднем мозгу двухолмие преобразуется в четверохолмие, в ростральных холмах которого находятся акустические центры.
Наблюдается дальнейшая дифференциация связей крыши среднего мозга с таламусом, который является как бы преддверия входа в кору всех восходящих сенсорных путей. В самом таламусе происходит дальнейшее обособление ядерных структур и установление между ними специализированных связей.
У млекопитающих развитие переднего мозга сопровождалось бурным ростом новой коры, находящейся в тесной функциональной связи с таламусом промежуточного мозга. В коре закладываются эфферентные пирамидные клетки, посылающие свои длинные аксоны к мотонейронам спинного мозга.
Корковая регуляция движений у млекопитающих приводит к развитию наиболее молодой части мозжечка — передней части задних долей полушарий, или неоцеребеллума. Неоцеребеллум приобретает двусторонние связи с новой корой.
Рост новой коры у млекопитающих происходит настолько интенсивно, что старая и древняя кора оттесняются в медиальном направлении к мозговой перегородке. Бурный рост коры компенсируется формированием складчатости.
У наиболее низко организованных однопроходных (утконос) на поверхности полушария закладываются первые две постоянные борозды, остальная же поверхность остается гладкой.
Как показали нейрофизиологические исследования, мозг однопроходных и сумчатых млекопитающих лишен еще соединяющего полушария мозолистого тела и характеризуется перекрытием сенсорных проекций в новой коре. Четкая локализация моторных, зрительных и слуховых проекций здесь отсутствует.
Мозг насекомоядных и грызунов характеризуется наличием мозолистого тела и дальнейшим увеличением общей площади новой коры, развитием борозд и извилин.
В процессе эволюции у хищных млекопитающих появляются теменные и лобные ассоциативные поля, ответственные за оценку биологически значимой информации, мотивацию поведения и программирование сложных поведенческих актов. Наблюдается дальнейшее развитие складчатости новой коры.
И наконец, приматы демонстрируют наиболее высокий уровень организации коры головного мозга. Кора приматов характеризуется шестислойностью, отсутствием перекрытия ассоциативных и проекционных зон. У приматов формируются связи между фронтальными и теменными ассоциативными полями и, таким образом, возникает целостная интегративная система больших полушарий.
Равновесные потенциалы ионов на мембране нейрона (уравнение Нернста). Качественный анализ зависимостей, которые это уравнение отражает. Вклад различных ионов в формирование мембранного потенциала нейрона.
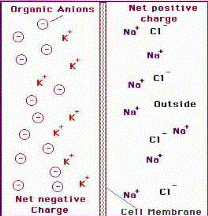
Концентрация ионов Na+ и Cl- выше во внеклеточном пространстве. Концентрация ионов K+ и органических анионов выше внутри клетки. В результате этого под действием электрохимических и электростатических сил возникает мембранный потенциал.
Электрические процессы в клетках обусловлены неравномерным распределением ионов по обе стороны клеточной мембраны.
В состоянии покоя нейрон изнутри заряжен отрицательно по отношению к своей внешней поверхности, т. е. обладает устойчивым мембранным потенциалом, или потенциалом покоя (ПП), который составляет в среднем -70 мВ. Концентрация основных ионов внутри и вне нервной клетки существенно различается; так, в аксоне кальмара ионов К+ в цитоплазме в 20—30 раз больше, чем снаружи; ионов Na+ и С1~ примерно в 10 раз меньше; Са2+ снаружи больше в 20—30 раз.
Мембранный потенциал. Вклад ионов К+
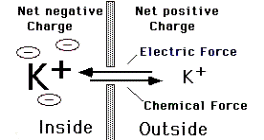 Мембрана нейрона обладает наибольшей проницаемостью для ионов К+. Вклад этих ионов в МП является наиболее значительным.
Мембрана нейрона обладает наибольшей проницаемостью для ионов К+. Вклад этих ионов в МП является наиболее значительным.
Электрохимическаясила (разница в концентрации иона внутри и внеклетки) приводит к выходу К+ из клетки, при этом из нее выносятся положительные заряды и возникает МП, накопление К+ на внешней поверхности мембраны препятствует дальнейшему выходу К+ из клетки – начинает действовать электростатическая сила в противоположном направлении. Потенциал, при котором наступает равновесие этих сил – калиевый равновесный потенциал.