 2015-04-20
2015-04-20 4831
4831Аня: Процесс суммации зависит от множества факторов, таких как электрохимические характеристики мембраны в месте суммации, количество и набор ионных каналов, степень их проводимости, количество и временная последовательность входных сигналов, «вес» каждого из каналов, т. е. коэффициенты и знаки, с которыми суммируются возбудительные и тормозные потенциалы в каждом месте мембраны и т. д.
Большое значение для результата суммации часто имеют геометрические параметры дендритных деревьев. Абсолютное большинство С. находится на различных веточках дендритных деревьев, длина ветвей которых может достигать миллиметров. Структура ветвлений и толщина отдельных веточек определяют входное сопротивление постсинаптических мембран в местах контактов и, т. о., вклад данного постсинаптического потенциала. В частности, более тонкие дендритные окончания обладают большим электрическим сопротивлением, в силу чего даже небольшие по амплитуде возбуждающие постсинаптические потенциалы вызывают на этих веточках полноценные потенциалы действия.
В итоге сложных и разнообразных процессов суммации возбуждающих и тормозных постсинаптических потенциалов, или, иначе говоря, в итоге процессов обработки информации в некотором участке нейронной сети суммарный постсинаптический потенциал снова преобразуется в последовательность нервных импульсов.
(то, что ниже пригодится и для вопросов 2 билетов 7 и 8)
Запуск импульсной активности в нервной системе осуществляют два основных фактора. Первый из них — стимулы, действующие на чувствительные клетки сенсорных систем и изменяющие проницаемость их мембраны. Это приводит к развитию особых рецепторных потенциалов и в итоге — к генерации ПД. Второй фактор — выделение медиатора из пресинаптического окончания. Попав в синаптическую щель, медиатор воздействует на постсинаптическую мембрану, возбуждая или тормозя следующий нейрон.
Процессы подобного возбуждения или торможения связаны с деятельностью еще одного типа ионных каналов — лиганд-зависимых (хемочувствительных). Они находятся на мембране, непосредственно окружающей синаптический контакт. Обычно они закрыты. Их открывание происходит лишь при появлении медиатора, несущего сигнал химического вещества (отсюда термин «хемочувствительные»).
Лиганд-зависимые каналы можно разделить на три основных класса: избирательно проницаемые по отношению к ионам Na+, ионам К+ и ионам С1-. Открывание первых из них приведет к входу в клетку ионов Na+ и деполяризации нейрона (рис. 3.14,а), во время которой разность потенциалов на мембране оказывается приближенной к порогу запуска ПД. В этот момент меньший, чем обычно, стимул может вызвать реакцию нейрона, т. е. нервная клетка находится в относительно возбужденном состоянии. В связи с этим локальная деполяризация мембраны под действием медиатора была названа возбуждающим постсинаптическим потенциалом (ВПСП).
Медиаторы, вызывающие ВПСП, отнесены к группе возбуждающих медиаторов.
Открывание хемочувствительных С1--каналов приводит к входу в клетку ионов хлора; открывание К+-каналов — к выходу ионов калия. В этих случаях возникает гиперполяризация и разность потенциалов на мембране нейрона увеличивается по абсолютной величине (рис. 3.14,б), поэтому для запуска ПД необходим больший, чем обычно, стимул. Следовательно, нервная клетка находится в относительно заторможенном состоянии. В связи с этим локальная гиперполяризация мембраны под действием медиатора была названа тормозным постсинаптическим потенциалом (ТПСП). Медиаторы, вызывающие ТПСП, отнесены к группе тормозных медиаторов.
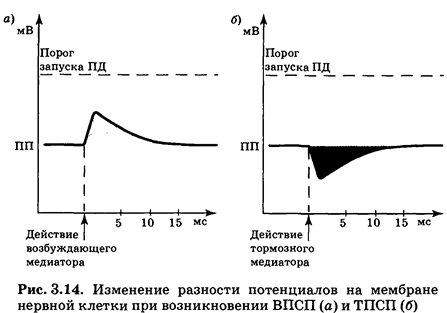
Усредненные параметры ВПСП и ТПСП весьма близки (рис. 3.14). Их длительность составляет обычно около 10мс (иногда 50—100 мс), что существенно больше, чем в случае ПД.
Амплитуда ВПСП и ТПСП определяется длительностью и крутизной наклона их первой фазы, которая зависит от количества и длительности существования медиатора в синаптической щели. Амплитуда одиночных постсинаптических потенциалов в ЦНС составляет 1—5 мВ. В крупном нервно-мышечном синапсе аналог ВПСП — потенциал концевой пластинки, может достигать 40 и более мВ.
При детальном анализе сигнала можно видеть, что первые фазы ВПСП и ТПСП имеют ступенчатый характер, т.е. нарастают дискретно, шагами (квантами). Такая дискретность связана с тем, что выброс медиатора в синаптическую щель так же идет квантами, где квант — одна везикула. В каждой везикуле содержится несколько тысяч молекул медиатора, и их воздействие на постсинаптическую мембрану вызывает сдвиг потенциала примерно на 0,1 мВ.
В подавляющем большинстве случаев (кроме потенциала концевой пластинки) одиночный ВПСП не способен запустить ПД, так как возбуждение, вызываемое медиатором, не дорастает до порогового уровня.
Для достижения порога запуска ПД необходима суммация (наложение) нескольких ВПСП.
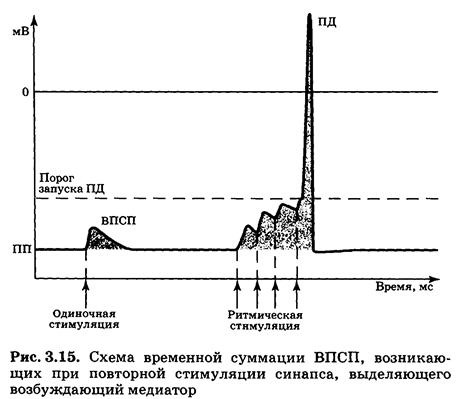
Выделяют два варианта суммации — временную и пространственную. Временная суммация — объединение эффектов стимулов, пришедших по одному «каналу» с большой частотой (рис. 3.15): если к еще не угасшему ВПСП присоединить второй, затем третий и т. д., — возникнет реальная возможность запустить ПД. Это означает, что сигнал, достигший синапса, достаточно интенсивен и «заслуживает» того, чтобы быть переданным дальше по сети нейронов.
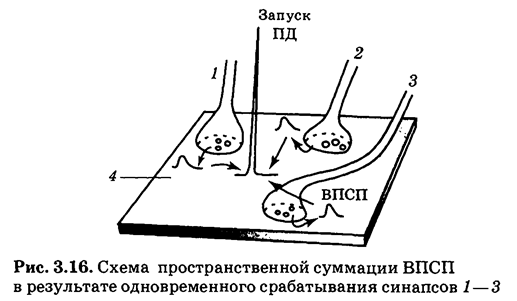
Пространственная суммация заключается в наложении друг на друга ВПСП соседних синапсов 1—3 в некоторой близлежащей точке постсинаптической мембраны 4 (рис. 3.16), обладающей потенциал-зависимыми ионными каналами. Схема пространственной суммации напоминает логическую ячейку по типу «И», т.е. результат положительный, если несколько условий будут выполнены (несколько входных сигналов одновременно достигнут нервной клетки).
В ходе деятельности нейронов эффекты пространственной и временной суммации объединяются, и чем больше синапсов участвуют в этом процессе (срабатывают относительно одномоментно), тем больше вероятность достичь порога запуска ПД. При этом часть синапсов может обладать тормозными свойствами и вызывать ТПСП, вычитающиеся из суммы возбуждающих влияний.
В результате в первом приближении условие запуска ПД в каждый момент времени можно определить следующим образом:
ПП + (сумма всех ВПСП) - (сумма всех ТПСП) > порог запуска ПД
Оценить вклад конкретных постсинаптических потенциалов в этот процесс непросто. Дело в том, что их влияние быстро затухает по мере удаления от места возникновения. Кроме того, затухание в отростках происходит быстрее, чем в теле нейрона, а также тем быстрее, чем тоньше отросток. И, наконец, мембрана нейрона в разных местах имеет разную возбудимость. Она максимальна в аксональном холмике (место отхождения аксона от тела) и в местах первого ветвления крупных дендритов. В итоге оказывается, что чем ближе конкретный синапс к этим точкам, тем больше его вклад в управление генерацией ПД. Одного ТПСП, возникшего рядом с аксональным холмиком, может оказаться достаточно для прекращения проведения сигнала (логическая ячейка по типу «НЕ»).
Процесс суммирования ВПСП и ТПСП, возникших в разных синапсах, является основной вычислительной операцией, протекающей на нейронах ЦНС. При ее реализации сигналы имеют возможность «подтвердить» свою значимость, могут объединиться с другими сигналами и сформировать некоторый «информационный образ», могут быть заблокированы (при наличии определенных условий — сигналов по тормозным каналам). В связи с этим способность ЦНС выполнять сложные вычислительные операции определяется не ее общим весом и даже не числом нейронов, а именно количеством синапсов. По мере развития мозг способен формировать дополнительные синапсы, увеличивая свои потенциальные возможности. Особенно интенсивно этот процесс идет в раннем детстве, когда нервная система настраивается на предстоящий уровень информационной нагрузки.
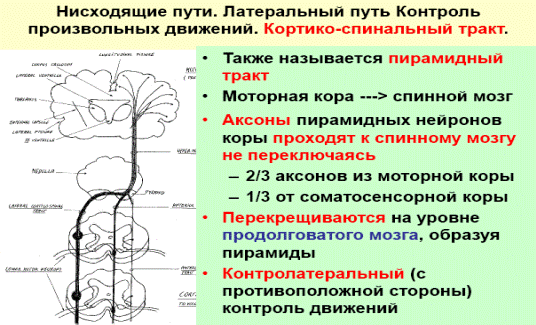
Пирамидный тракт описать по следующим параметрам: источник (структура головного мозга), какую эфферентную функцию обеспечивают, переключения (включая место перекреста, если он есть), конечный пункт
АНЯ: Нисходящий, идет от клеток Бенца в 5 слое коры полушарий головного мозга(двигательная зона), проходит через подкорковое белое вещество, задний отдел внутренней капсулы, ножки мозга, основание варолиева моста и пирамиды продолговатого мозга. На границе продолговатого и спинного мозга большинство волокон кортикоспинального пути переходят на противоположную сторону и далее следуют в составе боковых канатиков спинного мозга. Меньшая часть волокон кортикоспинального пути (от 10 до 30%) не перекрещивается и входит в передние канатики спинного мозга, осуществляет контроль за деятельностью дистальных участков конечностей, а также участвует в регуляции проведения импульсов от скелетной мускулатуры к высшим мозговым центрам.
Шеппард, стр 109

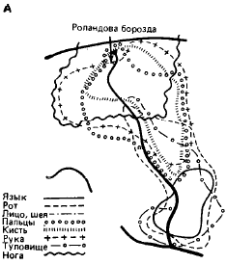

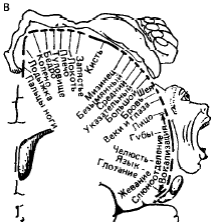







ЭКЗАМЕНАЦИОННЫЙ БИЛЕТ №10 по Физиологии ЦНС








