 2015-04-20
2015-04-20 2813
2813При поступлении потенциала действия к пресинаптическому окончанию происходит деполяризация пресинаптической мембраны и активация потенциал-зависимых кальциевых каналов. Кальций начинает поступать внутрь пресинаптического окончания и вызывает экзоцитоз везикул, наполненных нейромедиатором. Нейромедиатор выбрасывается в синаптическую щель и диффундирует к постсинаптической мембране. На поверхности постсинаптической мембраны медиатор связывается со специфическими рецепторами (лиганд-зависимыми каналами) и вызывает их открытие.
Различают следующие ПСП:
- Спонтанные и миниатюрные ПСП
- Потенциал концевой пластинки
- Вызванные ПСП
Потенциал действия (ПД). Все раздражители, действующие на клетку, вызывают в первую очередь снижение ПП(потенциал покоя); когда оно достигает критического значения (порога), возникает активный распространяющийся ответ - ПД. Во время восходящей фазы ПД кратковременно извращается потенциал на мембране: её внутренняя сторона, заряженная в покое электроотрицательно, приобретает в это время положительный потенциал. Достигнув вершины, ПД начинает падать (нисходящая фаза ПД), и потенциал на мембране возвращается к уровню, близкому к исходному ПП. Полное восстановление ПП происходит только после окончания колебаний потенциала - деполяризации или гиперполяризации, длительность которых обычно значительно превосходит продолжительность пика ПД. Согласно мембранной теории, деполяризация мембраны, вызванная действием раздражителя, приводит к усилению потока Na+ внутрь клетки, что уменьшает отрицательный потенциал внутренней стороны мембраны - усиливает её деполяризацию. Это, в свою очередь, вызывает дальнейшее повышение проницаемости для Na+ и новое усиление деполяризации и т.д. В результате такого взрывного кругового процесса, т. н. регенеративной деполяризации, происходит извращение мембранного потенциала, характерное для ПД. Повышение проницаемости для Na+ очень кратковременно и сменяется её падением, а следовательно, уменьшением потока Na+ внутрь клетки. Проницаемость для К+, в отличие от проницаемости для Na+, продолжает увеличиваться, что приводит к усилению потока К+ из клетки. В результате этих изменений ПД начинает падать, что ведёт к восстановлению ПП. Таков механизм генерации ПД в большинстве возбудимых тканей. Существуют, однако, клетки (мышечные волокна ракообразных, нервные клетки у ряда моллюсков, некоторые растительные клетки), у которых восходящая фаза ПД обусловлена повышением проницаемости мембраны не для ионов Na+, а для ионов Ca+. Своеобразен также механизм генерации ПД в мышечных волокнах сердца, для которых характерно длительное плато на нисходящей фазе ПД. Неравенство концентраций ионов К+ и Na+ (или Ca+) внутри и снаружи клетки (волокна) поддерживается специальным механизмом (т. н. "натриевым насосом"), выталкивающим ионы Na+ из клетки и нагнетающим ионы К+ в протоплазму, требующим затраты энергии, которая черпается клеткой в процессах обмена веществ.
Амплитуда ПД большинства нервных и мышечных волокон примерно одинакова: 110-120 мв. Длительность ПД варьирует в широких пределах: у теплокровных животных длительность ПД нервных волокон, наиболее быстро проводящих возбуждение, - 0,3-0,4 мсек, у волокон же мышц сердца - 50-600 мсек. В растительных клетках пресноводной водоросли хара ПД продолжается около 20 сек. Характерной особенностью ПД, отличающей его от других форм ответа клетки на раздражение, является то, что он подчиняется правилу "всё или ничего", т. е. возникает только при достижении раздражителем некоторого порогового значения, и дальнейшее увеличение интенсивности раздражителя уже не сказывается ни на амплитуде, ни на продолжительности ПД. Потенциал действия - один из важнейших компонентов процесса возбуждения. В нервных волокнах он обеспечивает проведение возбуждения от чувствительных окончаний (рецепторов) к телу нервной клетки и от неё - к синаптическим окончаниям, расположенным на различных нервных, мышечных или железистых клетках. Поступая в эффекторные окончания, ПД вызывает выделение (секрецию) определённой порции специфических химических веществ, т. н. медиаторов, оказывающих возбуждающее или тормозящее влияние на соответствующие клетки. В мышечных волокнах распространяющийся ПД вызывает цепь физико-химических реакций, лежащих в основе процесса сокращения мышц. Проведение ПД вдоль нервных и мышечных волокон осуществляется т. н. локальными токами, или токами действия, возникающими между возбуждённым (деполяризованным) и соседними с ним покоящимися участками мембраны. Токи действия регистрируются обычными внеклеточными электродами; при этом кривая имеет двухфазный характер: первая фаза соответствует приходу ПД под ближний электрод, вторая - под дальний электрод.
Постсинаптические потенциалы (ПСП) возникают в участках мембраны нервных или мышечных клеток, непосредственно граничащих с синаптическими окончаниями. Они имеют амплитуду порядка нескольких мв и длительность 10-15 мсек. ПСП подразделяются на возбуждающие (ВПСП) и тормозные (ТПСП). ВПСП представляют собой местную деполяризацию постсинаптической мембраны, обусловленную действием соответствующего медиатора (например, ацетилхолина в нервно-мышечном соединении). При достижении ВПСП некоторого порогового (критического) значения в клетке возникает распространяющийся ПД. ТПСП выражается местной гиперполяризацией мембраны, обусловленной действием тормозного медиатора. В отличие от ПД, амплитуда ПСП постепенно увеличивается с увеличением количества выделившегося из нервного окончания медиатора. ВПСП и ТПСП суммируются друг с другом при одновременном или последовательном поступлении нервных импульсов к окончаниям, расположенным на мембране одной и той же клетки.
Экстрапирамидную систему описать по следующим параметрам: какие тракты входят, источник (структура головного мозга), какую эфферентную функцию обеспечивают, переключения (включая место перекреста, если он есть), конечный пункт
Аня: Экстрапирамидная система состоит из следующих структур головного мозга:
· базальные ганглии
· красное ядро
· интерстициальное ядро
· тектум
· чёрная субстанция ретикулярная формация моста и продолговатого мозга
· ядра вестибулярного комплекса
· мозжечок
· премоторная область коры
· полосатое тело
Корковые экстрапирамидные пути слагаются из нервных волокон, идущих от клеток корковых двигательных центров к образованиям экстрапирамидной системы. Здесь можно выделить следующие пути: корково-таламические, корково-гипоталамические, корково-мостовые, корково-красноядерные и корково-покрышечные.
Например, кора полушарий большого мозга осуществляет управление функциями мозжечка, участвующего в координации движений, через мост по корково-мостомозжечковому пути. Их два: лобно-мостомозжечковый и затылочно-височно-мостомозжечковый. Они проходят из коры головного мозга к собственным ядрам моста и от них к коре мозжечка противоположной стороны.
Стриопаллидарные пути представлены нейритами клеток, залегающими в подкорковых базальных ядрах (в полосатом теле – в хвостатом ядре, бледном шаре, скорлупе); эти пути идут к ядрам таламуса, гипоталамуса, красного ядра, чёрного вещества. Анатомически указанные нейриты образуют три основных эфферентных пучка: чечевицеобразная петля, чечевицеобразный пучок и субталамический пучок.
Трункоспинальные пути образуются нервными проводниками, идущими от ядер среднего, промежуточного и продолговатого мозга к двигательным ядрам спинного мозга и черепных нервов, в составе следующих анатомически обособленных двигательных путей.
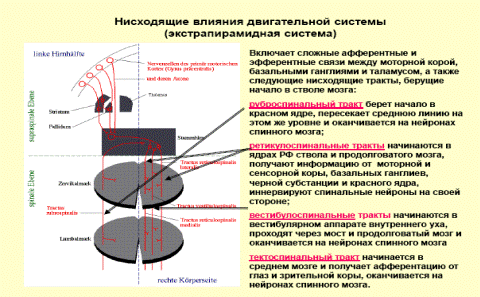
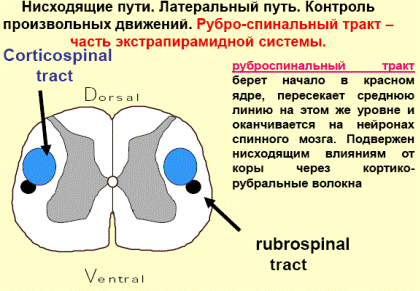
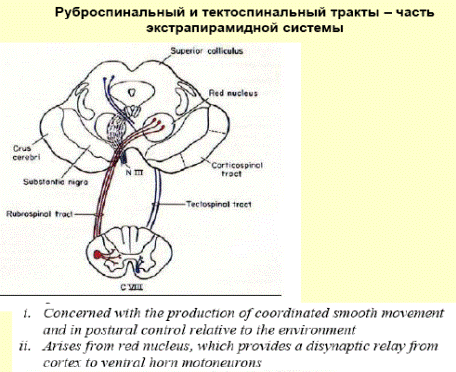
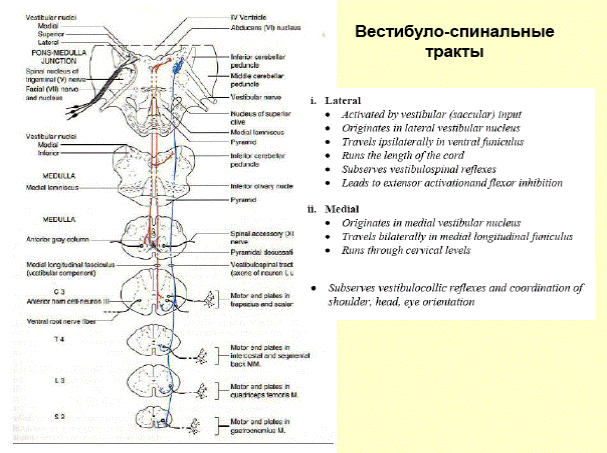
Одной из функций красного ядра является поддержание мышечного тонуса, необходимого для непроизвольного удержания тела в равновесии. От красного ядра нервные импульсы направляются в двигательные ядра передних рогов спинного мозга (красноядерно-спинальный путь). Он начинается из среднего мозга (от красного ядра), спускается по боковому канатику противоположной стороны спинного мозга и оканчивается на двигательных нейронах передних рогов. Этот путь несёт непроизвольные двигательные импульсы, он имеет важное значение для экстрапирамидного обеспечения движений.
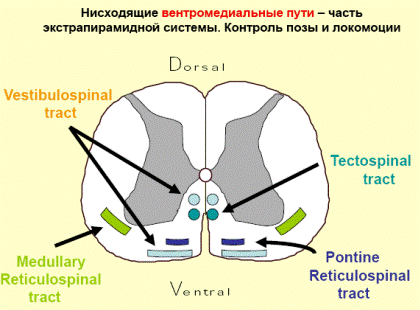
В осуществлении координации движений человека при нарушении равновесия важную роль играет преддверно-спинномозговой путь, который лежит между передним и боковым канатиками и соединяет вестибулярные ядра ромбовидной ямки с передними рогами спинного мозга. Первый нейрон этого пути залегает в ядрах 8-ой пары черепных нервов. Эти ядра соединены с мозжечком и, посредством заднего продольного пучка, с двигательными ядрами 3-ей, 4-ой и 6-ой пар черепных нервов. Это обеспечивает сохранение положения глазного яблока при движениях головы и шеи. Аксоны вторых нейронов преддверно-спинномозгового пути спускаются вниз в составе переднего канатика спинного мозга и заканчиваются синапсами на двигательных клетках передних рогов спинного мозга. Нейроны ретикулярной формации обеспечивают связь преддверно-спинномозгового пути с базальными ядрами.
Покрышечно-спинномозговой путь начинается от ядер покрышки четверохолмия и заканчивается у клеток передних рогов шейных сегментов, устанавливает связи экстрапирамидной системы, а также подкорковых центров зрения и слуха с шейной мускулатурой; таким образом он связан со слуховыми и зрительными восприятиями.
Оливоспинномозговой путь присутствует в шейных сегментах спинного мозга; начинается от нейронов оливы и заканчивается на клетках передних рогов.
Спинно-ретикулярный путь идёт от ретикулярной формации ствола головного мозга к мотонейронам спинного мозга, осуществляет большое влияние на функции спинного мозга.
Задний продольный пучок начинается от ядра Даркшевича и заканчивается посегментно у мотонейронов спинного мозга. Имеет связи со всеми ядрами глазодвигательных нервов и вестибулярного нерва. Обеспечивает одновременность поворота глазных яблок и головы, содружественность движений глазных яблок. Этот пучок называют также пучком Шютца.
Медиальный продольный пучок лежит в переднем канатике и состоит из нисходящих и восходящих волокон; берёт начало и оканчивается на ядрах ствола мозга и на клетках передних рогов; иннервирует мышцы шеи. Пучок представляет собой очень древнюю систему волокон, которая у низших позвоночных служит важнейшим ассоциационным путём головного мозга.
ЭКЗАМЕНАЦИОННЫЙ БИЛЕТ №11 по Физиологии ЦНС
Разнообразие нейронов. Связь строения и функции. Опишите основные классы нейронов по строению и укажите, в каком отделе нервной системы (сенсорном, моторном, центральном) они представлены. Приведите примеры.
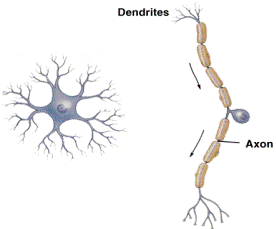
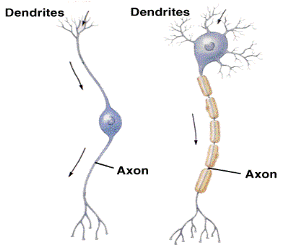
Анаксонные униполярные биполярные мультиполярные
Амакриновые (unipolar, (bipolar) (multipolar)
(anaxonic) pseudounipolar)








