По биологическим функциям гормоны можно разделить на несколько групп.
1. Первый уровень — ЦНС. Нервные клетки получают сигналы из внешней и внутренней среды, преобразуют их в форму нервного импульса и передают через синапсы, используя химические сигналы — медиаторы. Медиаторы вызывают изменения метаболизма в эффекторных клетках.
2. Второй уровень — эндокринная система. Включает гипоталамус, гипофиз, периферические эндокринные железы, а также отдельные клетки (АПУД система), синтезирующие под влиянием соответствующего стимула гормоны, которые через кровь действуют на ткани-мишени.
3. Третий уровень — внутриклеточный. На метаболические процессы в клетке влияют субстраты и продукты обмена веществ, а также тканевые гормоны (аутокринно).
На основании иерархических уровней существует несколько осей регуляции:
1. гипоталамо-гипофизарно-надпочечниковая ось;
2. гипоталамо-гипофизарно-тиреоидная ось;
3. симпато-адреналовая ось;
Регуляции обмена веществ в тканях мишенях осуществляется через изменения активности и количества ферментов, изменения скорости транспорта веществ через мембраны клеток.
Принципы организации нейроэндокринной системы
В основе работы нейроэндокринной системы лежит принцип прямой, обратной, положительной и отрицательной связи.
Принцип прямой положительной связи – активация текущего звена системы приводит к активации следующего звена системы, распространению сигнала в сторону клеток-мишеней и возникновению метаболических или физиологических изменений.
Принцип прямой отрицательной связи – активация текущего звена системы приводит к подавлению следующего звена системы и прекращению распространения сигнала в сторону клеток-мишеней.
Принцип обратной отрицательной связи – активация текущего звена системы вызывает подавление предыдущего звена системы и прекращение его стимулирующего влияния на текущую систему.
Принципы прямой положительной и обратной отрицательной связи являются основой для поддержания гомеостаза.
Принцип обратной положительной связи – активация текущего звена системы вызывает стимуляцию предыдущего звена системы. Основа циклических процессов.
СХЕМА!
Действие гормона. Рецепторы к СТГ есть в печени, адипоцитах, яичках, желтом теле, скелетных мышцах, хрящевой ткани, мозге, легких, поджелудочной железе, кишечнике, сердце, почках, лимфоцитах. Действует через инозитолтрифосфатную систему.
Кратковременных эффекты СТГ: В адипоцитах активируется потребление глюкозы и липогенез, что снижает уровень глюкозы в крови.
Долговременные эффекты СТГ: В адипоцитах усиливается липолиз, повышается в крови концентрация жирных кислот (при дефиците инсулина повышается в крови концентрация кетоновых тел). В печени ускоряется глюконеогенез, а в адипоцитах, мышцах снижается потребление глюкозы, что вызывает гипергликемию.
Основное действие СТГ направлено на регуляцию обмена белка, процесса роста и развития организма. СТГ усиливает транспорт АК в мышцы, синтез белка в костях, хрящах, мышцах, печени и т.д. СТГ увеличивает количество РНК, ДНК, общее число клеток. СТГ увеличивает рост костей, соединительной ткани, мышц и внутренних органов.
Инактивация. Катаболизм СТГ происходит гидролизов до АК в тканях мишенях. Т½ СТГ=50 мин.
ГОРМОНЫ КОРЫ НАДПОЧЕЧНИКОВ
Гипоталамо-гипофизарно-надпочечниковая ось
В коре надпочечников синтезируется более 40 различных стероидов, различающихся по структуре и биологической активности. Кортикостероиды в зависимости от их преобладающего действия делят на 3 основные класса: глюкокортикоиды, минералокортикоиды и андрогены.
1. Синтез кортикостероидов
Общим предшественником кортикостероидов служит ХС, который поступает из крови в составе ЛПНП или синтезируется из ацетил-КоА и хранится в цитоплазме в виде эфиров.
ФОРМУЛА!
Расширяют сосуды, увеличивают их проницаемость. Вызывают сокращение бронхов. Основные компоненты «медленно реагирующей субстанции» анафилаксии.
Лейкоциты, альвеолярные макрофаги
LXA4 липоксин
Стимулирует хемотаксис и образование супероксид аниона в лейкоцитах
Лейкоциты
гранулоцитомакрофагальный колониестимулирующий фактор, макрофагальный колониестимулирующий фактор, лейкемический ингибиторный фактор, онкостатин М, фактор стволовых клеток и др.
Активированные лимфоциты и другие иммунокомпетентные клетки секретируют как факторы роста (нервный, эпидермальный, b-трансформирующий фактор роста, соматомедины, или инсулиноподобныый фактор роста 1 и 2 (ИФР 1 и 2), так и различные полипептидные гормоны (АКТГ, ТТГ, ЛГ, ФСГ, СТГ, пролактин, хорионический гонадотропин, соматостатин, ВИП, окситоцин, вазопрессин, метэнкефалин, кортиколиберин, соматолиберин, вещество Р и др.).
В тимусе (вилочковой железе) синтезируются тимические гормоны: тимозины, тимический гуморальный фактор, тимопоэтин, тимулин, тимический фактор Х, тимостимулин. Кроме того, в тимусе вырабатываются перечисленные выше лимфокины (интерлейкин 1, 2, 4, 6, 7, фактор некроза опухолей и др.), нейропептиды (нейротензин, вещество Р, ВИП, холецистокинин, соматостатин, окситоцин, вазопрессин, нейротензины, метэнкефалин, АКТГ, предсердный натрийуретический пептид).
· Цитокины и тимические гормоны осуществляют свое действие аутокринным или паракринным путем, влияя на дифференцировку Т-клеток, увеличивая количество Т-супрессоров, восстанавливая реактивность Т-клеток, влияя на гемопоэтические клетки.
· Цитокины способны влиять на работу гипофиза, гипоталамуса и периферических эндокринных желез.
· Гормоны и их рецепторы часто служат объектом аутоиммунного ответа, а при патологии мишенью аутоаллергии.
ЭЙКОЗАНОИДЫ
Эйкозаноиды – БАВ, образуются из полиеновых ЖК с 20 атомами С (арахидоновая, эйкозапентаеновая, эйкозатриеновая). Эйкозаноиды являются тканевыми гормонами (аутокринный и паракринный эффект), с коротким периодом полураспада (секунды - минуты). Концентрация эйкозаноидов в крови низкая. Системное действие оказывают при некоторых патологиях, когда их концентрация в крови заметно повышается.
Схема образования эйкозаноидов
Полиеновые ЖК с 20 атомами С поступают в организм с пищей или синтезируются из эсенциальных полиеновых ЖК с 18 атомами С.
После выделения арахидоновой кислоты из глицерофосфолипида она выходит в цитозоль и превращается 2 путями в эйкозаноиды: 1 путь циклооксигеназный дает простагландины, простациклины и тромбоксаны; 2 путь липоксигеназный дает лейкотриены, липоксины и др. эйкозаноиды.
Эйкозаноид PGE1: PG – простагландин, Е – заместитель в пятичленном кольце эйкозаноида, 1 – число двойных связей в боковых цепях эйкозаноида.
PG – простагландины, имеют 2 кольца в структуре (пятичленное и эндопероксидное).
ФОРМУЛА!
· Йодтиронины участвуют в регуляции многих процессов метаболизма, развития и клеточной дифференцировки.
· При физиологической концентрации йодтиронины ускоряют белковый синтез, стимулируют процессы роста и клеточной дифференцировки, ускоряют транскрипцию гена гормона роста.
· В печени йодтиронины ускоряют гликолиз, синтез холестерола и синтез жёлчных кислот. В печени и жировой ткани Т3 повышает чувствительность клеток к действию адреналина и косвенно стимулирует липолиз в жировой ткани и мобилизацию гликогена в печени. Т3 увеличивает в мышцах потребление глюкозы, стимулирует синтез белков и увеличение мышечной массы, повышает чувствительность мышечных клеток к действию адреналина.
· Йодтиронины стимулируют работу Na+,K+-ATФазы, повышают поглощение клетками кислорода (кроме мозга, РЭС и гонад).
· Йодтиронины участвуют в формировании ответной реакции на охлаждение увеличением теплопродукции, повышая чувствительность симпатической нервной системы к норадреналину и стимулируя секрецию норадреналина.
· Очень высокие концентрации Т3 тормозят синтез белков и стимулируют катаболические процессы.
6. Инактивация йодтиронинов осуществляется в периферических тканях в результате дейодирования Т4 до «реверсивной» Т3 по 5, полного дейодирования, дезаминирования или декарбоксилирования. Йодированные продукты катаболизма йодтиронинов конъюгируют в печени с глюкуроновой или серной кислотами, секретируются с жёлчью, в кишечнике вновь всасываются, дейодируются в почках и выделяются с мочой. Для Т4 Т½ =7 дней, для Т3 Т½ =1-1,5 дня.
Регуляция синтеза и секреции йодтиронинов
Синтез и секреция йодтиронинов регулируется гипоталамо-гипофизарной системой.
ФОРМУЛА!
4. Транспорт
Тестостерон транспортируется в крови в основном в комплексе с альбумином (40%) и специфически связывающим половые гормоны β-глобулином (называемым секс-гормонсвязывающим глобулином, СГСГ). Лишь 2% тестостерона находиться в крови в свободном виде.
Действие гормонов. Тестостерона регулирует синтез белков в эмбриональных вольфовых структурах, сперматогониях, мышцах, костях, почках и мозге. Действие андрогенов в онтогенезе различно.
У эмбриона под действием андрогенов из вольфова протока образуются придаток яичка (эпидидимис), семявыносящий проток и семенной пузырёк, происходит маскулинизация мозга.
В препубертатный период андрогены оказывают мощное анаболическое действие, стимулируют клеточное деление. Они скачкообразно увеличивают линейный размер тела, увеличивают скелетные мышцы, рост костей, но одновременно способствуют и остановке роста, так как стимулируют сращение эпифизов длинных костей с их стволами. Андрогены вызывают изменение структуры кожи и волос, снижение тембра голоса вследствие утолщения голосовых связок и увеличения объёма гортани, стимулируют секрецию сальных желёз.
ЖЕНСКИЕ ПОЛОВЫЕ ГОРМОНЫ
В яичниках синтезируются женские половые гормоны — эстрогены и прогестины, среди которых наиболее активны 17β-эстрадиол и прогестерон.
Синтез.
1. Превращение ХС в прегненалон катализирует в митохондриях клеток теки фолликулов холестеролдесмолаза (Р450), ее активирует через аденилатциклазную систему ЛГ. В клетках теки фолликулов синтезируются андрогены (андростендион). В клетках теки синтезируется очень небольшое количество эстрогенов.
2. В клетках гранулёзы (яичников) андрогены с участием Р450-оксидазы (требует О2 и НАДФH2) ароматизируются в эстрадиол. Р450-оксидазу в клетках гранулёзы активирует ФСГ. В клетках гранулёзы может синтезироваться менее активный эстроген— эстрон.
Значительная часть андрогенов ароматизируется в эстрогены на периферии: в жёлтом теле, фетоплацентарном комплексе (во время беременности), корой надпочечников, в жировых клетках, печени, коже и других тканях.
Малоактивный эстриол образуется из эстрона в крови.
Прогестерон выделяется главным образом жёлтым телом во время менструации (в лютеиновую фазу) и фетоплацентарным комплексом во время беременности. В небольших количествах он вырабатывается у женщин и мужчин корой надпочечников.
В фолликулярной фазе менструального цикла концентрация прогестерона в плазме обычно не превышает 5 нмоль/л, а в лютеиновой фазе увеличивается до 40—50 нмоль/л.
СекрецияРегуляция секреции
1. Из гипоталамуса импульсно секретируется гонадотропин-рилизинг-гормон (декапептид).
2. Гондотропный гормон импульсно стимулирует в гипофизе синтез и секрецию фолликулостимулирующего (ФСГ) и лютеинизирующего (ЛГ) гормонов (гонадотропные гормоны гипофиза, гликопротеины, около 30 кД).
3. ЛГ у женщин стимулирует продукцию андрогенов клетками теки, образование прогестерона клетками жёлтого тела.
· ФСГ ускоряет развитие фолликулов в яичниках, стимулирует ароматизацию андрогенов с образованием эстрогенов.
Превращение прегненолона в тестостерон катализируется пятью микросомальными ферментами и может протекать двумя путями: через образование дегидроэпиандростерона или через образование прогестерона (что, по-видимому, преобладает в семенниках человека).
Дигидротестостерон. В семенных канальцах, предстательной железе, коже, наружных половых органах из 4% тестостерона под действием цитоплазматической НАДФН2-зависимой 5-α-редуктазы образуется более активный андроген — дигидротестостерон (400 мкг/сут).
В некоторых периферических тканях небольшое количество тестостерона превращается в эстрадиол. В качестве побочных продуктов клетки Лейдига также постоянно секретируют эстрадиол и прогестерон, хотя роль этих гормонов в развитии и поддержании функций размножения и формирования полового поведения у мужчин до настоящего времени не выяснена.
ФОРМУЛА!
сознания. Такое состояние возникает вследствие нарушения обмена электролитов, которое приводит к потере ионов Na+ и С1- с мочой, обезвоживанию за счёт потери внеклеточной жидкости, повышению уровня К+ в сыворотке крови, в межклеточной жидкости и клетках, в результате чего может нарушаться сократительная способность миокарда. Изменение углеводного обмена проявляется в снижении уровня сахара в крови, уменьшении запаса гликогена в печени и скелетных мышцах.
Первичная недостаточность надпочечников (болезнь Аддисона)
Причина: развивается в результате поражения коры надпочечников туберкулёзным или аутоиммунным процессом.
Основные клинические проявления выражаются в снижении массы тела, общей слабости, снижении аппетита, тошноте, рвоте, снижении АД и типичной для первичной надпочечниковой недостаточности гиперпигментации кожи («бронзовая болезнь»). Причина гиперпигментации — повышение продукции ПОМК — предшественника АКТГ и меланоцитстимулирующего гормона.
Вторичная недостаточность надпочечников
Причина: может развиться при дефиците АКТГ, вследствие опухоли или инфекционного поражения гипофиза. В отличие от болезни Аддисона, отсутствует гиперпигментация.
Врождённая гиперплазия надпочечников
Причина: нарушается синтез кортизола. В 95% случаев при этой патологии обнаруживается дефект 21-гидроксилазы (реже 11-гидроксилазы). Снижение продукции кортизола сопровождается увеличением секреции АКТГ, накоплением промежуточных продуктов синтеза кортикостероидов, в частности, предшественников андрогенов. Избыток андрогенов ведёт к усилению роста тела, раннему половому созреванию у мальчиков и развитию мужских половых признаков у девочек (адреногенитальный синдром). При частичной недостаточности 21-гидроксилазы у женщин может нарушаться менструальный цикл.
Гиперпродукция глюкокортикоидов (гиперкортипизм)
Причина: может быть следствием повышения уровня АКТГ при опухолях гипофиза болезньИценко-Кушинга) и опухолях других клеток (бронхов, тимуса, поджелудочной железы), вырабатывающих кортикотропинподобные вещества, или избыточного синтеза кортизола при гормонально-активных опухолях коры надпочечников (синдром Иценко-Кушинга).
Основные проявления: гипергликемия и снижение толерантности к глюкозе, обусловленные стимуляцией глюконеогенеза («стероидный диабет»), усиление катаболизма белков, уменьшение мышечной массы, истончение кожи, остеопороз, инволюция лимфоидной ткани. Характерно своеобразное перераспределение отложений жира («лунообразное лицо», выступающий живот). Гипернатриемия, гипертензия, гипокалиемия обусловлены некоторой минералокортикоидной активностью кортизола, которая проявляется при его избытке.
ПОЛОВЫЕ ГОРМОНЫ
Репродуктивные функции организма регулируются половыми гормонами: у мужчин — тестостероном, у женщин — эстрогенами и прогестинами.
МУЖСКИЕ ПОЛОВЫЕ ГОРМОНЫ
Мужские половые гормоны вырабатываются в основном в мужских половых железах — интерстициальных клетках Лейдига семенников (95%), небольшое количество андрогенов образуется в коре надпочечников.
1. Синтез андрогенов
Самый активный минералокортикоид — альдостерон.
Б. Синтез альдостерона
1. В цитозоле прегненолон (из митохондрий) под действием 3-β-гидроксистероиддегидрогеназы превращается в прогестерон.
2. В мембране ЭР 21-гидроксилаза гидроксилирует прогестерон в 11-дезоксикортикостерон.
3. В мембране ЭР 11-гидроксилаза гидроксилирует 11-дезоксикортикостерон в кортикостерон, обладающего слабовыраженной глюкокортикоидной и минералокортикоидной активностью.
4. В митохондриях 18-гидроксилаза гидроксилирует кортикостерон в альдостерон.
В. Синтез андрогенов и их предшественников
В коре надпочечников образуются предшественники андрогенов дегидроэпиандростерон (ДЭА) (наиболее активный) и андростендион (слабый), и в малых количествах гормоны - тестостерон и эстрадиол. В норме продукция надпочечниками активных андрогенов не играет существенной роли.
Предшественники андрогенов превращаются в активные андрогены в гонадах.
2. Секрециякортикостероидов происходит пассивно.
3. Регуляция. Синтез и секрецию кортизола стимулирует стресс, травма, инфекция, понижение концентрации глюкозы в крови. Повышение концентрации кортизола подавляет синтез кортиколиберина и АКТГ по механизму отрицательной обратной связи.
Главный стимул для синтеза и секреции альдостерона является ангиотензин II. Секрецию альдостерона также стимулируют низкая концентрация Na+, высокая концентрацией К+ в плазме крови, АКТГ, простагландины.
Базальный уровень кортикостероидов претерпевает в течение суток характерные колебания, связанные с центральным ритмоводителем организма, находящемся в гипоталамусе, и с системой регуляции цикла сон — бодрствование. Пик секреции кортикостероидов у человека приурочен к ранним утренним часам, когда наблюдается максимальное «давление» парадоксального сна — той стадии ночного сна, когда отмечаются высокий уровень активности головного мозга и эмоционально окрашенные сновидения.
4. Транспорт. Кортизол в плазме крови транспортируется в комплексе с α-глобулином транскортином и в небольшом количестве, около 8%, в свободной форме. Синтез транскортина протекает в печени и стимулируется эстрогенами.
Альдостерон не имеет специфического транспортного белка, транспортируется альбумином. Небольшое количество альдостерона находится в плазме крови в свободном виде.
6. Инактивация. Катаболизм кортикостероидов происходит в основном в печени. Кортикостероиды в реакциях гидроксилирования, окисления и восстановления образуют 17-кетостероиды, большая часть которых конъюгирует с глюкуроновой и серной кислотами. Затем 17-кетостероиды (кроме кортикостерона и альдостерона) выводятся с мочой. Т½ кортизола =1,5-2 ч.
Действие гормонов. Кортикоиды регулируют метаболизм, в основном меняя синтез ферментов (регуляция транскрипции генов в клетках-мишенях).
Глюкокортикоиды, играют важную роль в развитии стрессовой реакции. Они оказывают разнообразные эффекты, но наиболее важный — стимуляция глюконеогенеза. Кортизол:
· стимулирует глюконеогенез в печени.
Структурная организация нейроэндокринной системы.
Этапы метаболизма гормонов
Основное свойство всех живых организмов – поддержание гомеостаза. Нарушение гомеостаза приводит к смерти. У человека в подержании гомеостаза участвуют 3 основные системы:
1. Центральная и периферическая нервные системы через нервные импульсы и нейромедиаторы регулируют физиологические функции и работу эндокринной системы;
2. Эндокринная система через гормоны эндокринных желез регулирует метаболические и физиологические процессы, пролиферацию, дифференцировку клеток и тканей;
3. Иммунная система через специфические белки цитокины и антитела защищает организм от внешних и внутренних патогенных факторов, регулирует иммунные и воспалительные реакции, пролиферацию, дифференцировку клеток, работу эндокринной системы;
Системы регуляции обмена веществ и функций организма образуют 3 иерархических уровня.
Концепция ткани мишени
В организме около 200 типов дифференцированных клеток, лишь некоторые из них продуцируют гормоны, но все являются мишенями для действия гормонов.
Ткань мишень – ткань, в которой гормон вызывает специфическую биохимическую или физиологическую реакцию. Клетки тканей мишеней для взаимодействия с гормоном синтезируют специальные рецепторы, количество и тип которых определяет интенсивность и характер ответа.
ГОРМОНЫ
· Гормоны – органические сигнальные молекулы беспроводного системного действия. Гормоны – органические вещества, синтезирующиеся в эндокринных железах, транспортируемые кровью и действующие на ткани мишени (гормоны щитовидной железы, надпочечников, поджелудочной железы и т.д). Всего известно более 100 гормонов.
· Гормоноподобные вещества - органические вещества, синтезирующиеся апудоцитами, транспортируемые кровью и действующие на ткани мишени. Апудоциты – это диффузные эндокриноциты (отдельные клетки, не оформленные в железу), они образуются из эктодермы, эндодермы или мезодермы. Апудоциты формируют АПУД систему (диффузную гормональную систему). Апудоциты находятся в ЖКТ, вилочковой железе, сердце, печени, почках, ЦНС, плаценте и коже. Часто гормоноподобные вещества имеют то же самое строение, что и истинные гормоны, нейромедиаторы. Например, в ЖКТ синтезируются вазоактивный интестинальный пептид (ВИП), холецистокинин, гастрин, нейротензин, мет-, лейэнкефалин и др..
· Тканевые гормоны - органические вещества, синтезирующиеся отдельными клетками, не транспортируемые кровью и действующие на ткани мишени.
Особенности действия гормонов:
1. Действуют в малых количествах (10-6-10-12 ммоль/л);
2. Существует абсолютная или высокая специфичность в действии гормонов.
3. Переносят только информацию. Не используются в энергетических и строительных целях;
4. Действуют опосредованно через рецепторы и внутриклеточные посредники (Са2+, цАМФ, цГМФ, ДАГ, ИФ3 и т.д.). Например, через аденилатциклазную, инозитолтрифосфатную системы;
5. Регулируют активность или количество ферментов;
6. Зависят от ЦНС;
7. Беспороговый принцип. Даже 1 молекула гормона способна оказать эффект;
8. Пермессивность действия. Конечный эффект - результат действия множества гормонов.
Гормоны могут оказывать как системное, так и местное действие.
Эндокринное (системное) действие гормонов (эндокринный эффект) реализуется, когда они транспортируются кровью и контактируют с органами и тканями всего организма. Характерно для истинных гормонов и гормоноподобных веществ.
Местное действие гормонов реализуется, когда они влияют на клетки, в которых были синтезированы (аутокринный эффект), или на соседние клетки (паракринный эффект). Характерно для тканевых гормонов, есть также у истинных гормонов и гормоноподобных веществ.
Классификация и номенклатура гормонов
Эта классификация условна, поскольку одни и те же гормоны могут выполнять разные функции. Например, адреналин участвует в регуляции обмена жиров и углеводов и, кроме этого, регулирует частоту сердечных сокращений, АД, сокращение гладких мышц. Кортизол не только стимулирует глюконеогенез, но и вызывает задержку NaCl.
 2015-04-30
2015-04-30 1120
1120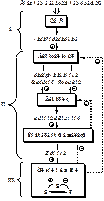






 8463
8463 7918
7918