 2015-05-10
2015-05-10 3242
3242После завершения репликации происходит метилирование нуклеотидных остатков вновь образованных цепей ДНК. Метальные группы присоединяются ко всем остаткам аденина в последовательности -GATC-, при этом образуется N6-метиладенин, а также возможны метилирование цитозина в последовательности -GC-и образование N5-метилцитозина. Количество метилированных оснований равно примерно 1-8%. Модификация происходит при участии ферментов, использующих в качестве источника метальных групп S-аденозилметионин (SAM). Присоединение метальных рупп к остаткам аденина и цитозина не нарушает комплементарности цепей.
Наличие метальных групп в цепях ДНК необходимо для формирования структуры хромосом, а также для регуляции транскрипции генов. В течение непродолжительного времени в молекуле ДНК последовательности -GATC-метилированы по аденину только в матричной, но не в новой цепи. Это различие используется ферментами репарации для исправления ошибок, которые могут возникать при репликации.
Транскрипция и трансляция.
Экспрессия гена — процесс реализации информации, закодированной в гене. Состоит из двух основных стадий — транскрипции и трансляции.
Транскрипция — синтез РНК на матрице ДНК. В ходе процесса образуются молекулы мРНК, служащие матрицей для синтеза белков, а также транспортные, рибосомальные и другие виды молекул РНК, выполняющие структурные, адапторные и каталитические функции.
Транскрипт — продукт транскрипции, т. е. РНК, синтезированная на данном участке ДНК-матрицы.
Промотор — регуляторный участок гена, к которому присоединяется РНК-полимераза с тем, чтобы начать транскрипцию.
Элонгация – удлинение цепи РНК за счет комплементарного присоединения новых нуклеотидов.
Терминатор – это участок, где прекращается дальнейший рост цепи РНК и происходит ее освобождение от матрицы ДНК.
Процессинг – совокупность событий, связанных с претрансляционным преобразованием первичного РНК-транскрипта.
Экзон — значащий участок гена, на котором записана информация о порядке аминокислот в молекуле белка. Сохраняется при сплайсинге.
Интрон — некодирующий участок гена, который переписывается на gРНК, а затем удаляется из нее при сплайсинге.
Сплайсинг — процесс формирования зрелой и-РНК путем удаления внутренних частей молекулы — интронов.
Энхансер — регуляторный участок ДНК, усиливающий транскрипцию с ближайшего к нему промотора.
Индуктор — фактор (вещество, свет, теплота и т.п.), вызывающий транскрипцию генов, находящихся в неактивном состоянии.
Этапы транскрипции:
Присоединение РНК-полимеразы
Инициация
Элонгация
Терминация
Синтез молекул РНК начинается в определённых последовательностях ДНК(промоторах) и завершается в терминирующих участках (сайты терминации). Участок ДНК, ограниченный промотором и сайтом терминации, представляет собой единицу транскрипции - транскриптон. В каждом транскриптоне присутствует неинформативная зона; она содержит специфические последовательности нуклеотидов, с которыми взаимодействуют регуляторные транскрипционные факторы.
Транскрипционые факторы - белки, взаимодействующие с определёнными регуляторными сайтами и ускоряющие или замедляющие процесс транскрипции. Соотношение информативной и неинформативной частей в транскриптонах эукариотов составляет в среднем 1:9 (у прокариотов 9:1).
Соседние транскриптоны могут быть отделены друг от друга нетранскрибируемыми участками ДНК. Разделение ДНК на множество транскриптонов позволяет осуществлять с разной активностью индивидуальное считывание (транскрипцию) разных генов.
Синтез цепи РНК идёт от 5'- к З'-концу, при этом матричная цепь ДНК всегда антипараллельна синтезируемой нуклеиновой кислоте.
Биосинтез РНК осуществляется ДНК-зависимыми РНК-полимеразами. В ядрах эукариотов обнаружены 3 специализированные РНК-полимеразы: РНК-полимераза I, синтезирующая пре-рРНК; РНК-полимераза II, ответственная за синтез пре-мРНК; РНК-полимераза III, синтезирующая пре-тРНК. РНК-полимеразы - олигомерные ферменты, состоящие из нескольких субъединиц - 2α, β, β', σ. Субъединица о (сигма) выполняет регуляторную функцию, это один из факторов инициации транскрипции, РНК-полимеразы I, II, III, узнающие разные промоторы, содержат разные по строению субъединицы σ.
Стадии транскрипции.
Инициация
Активация промотора происходит с помощью большого белка - ТАТА-фактора, называемого так потому, что он взаимодействует со специфической последовательностью нуклеотидов промотора - ТАТААА- (ТАТА-бокс)
Присоединение ТАТА-фактора облегчает взаимодействие промотора с РНК-полимеразой. Факторы инициации вызывают изменение конформации РНК-полимеразы и обеспечивают раскручивание примерно одного витка спирали ДНК, т.е. образуется транскрипционная вилка.
Элонгация
Факторы элонгации повышают активность РНК-полимеразы и облегчают расхождение цепей ДНК. Синтез молекулы РНК идёт от 5'- к З'-концу комплементарно матричной цепи ДНК. На стадии элонгации, в области транскрипционной вилки, одновременно разделены примерно 18 нуклеотидных пар ДНК. Растущий конец цепи РНК образует временную гибридную спираль, около 12 пар нуклеотидных остатков, с матричной цепью ДНК. По мере продвижения РНК-полимеразы по матрице в направлении от 3'- к 5'-концу впереди неё происходит расхождение, а позади - восстановление двойной спирали ДНК.
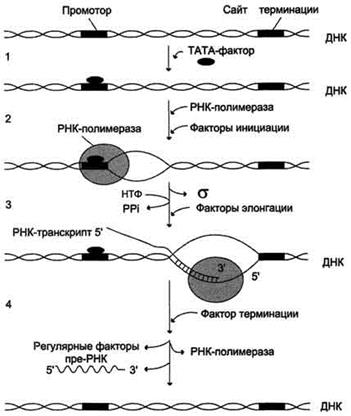
Рис. 4-30. Стадии транскрипции. 1 - присоединение ТАТА-фактора к промотору. Чтобы промотор был узнан РНК-полимера-зой, необходимо образование транскрипционного комплекса ТАТА-фактор/ТАТА-бокс (промотор). ТАТА-фактор остаётся связанным с ТАТА-боксом во время транскрипции, это облегчает использование промотора многими молекулами РНК-полимеразы; 2 - образование транскрипционной вилки; 3 - элонгация; 4.- терминация.
Терминация
Раскручивание двойной спирали ДНК в области сайта терминации делает его доступным для фактора терминации. Завершается синтез РНК в строго определенных участках матрицы - терминаторах (сайты терминации). Фактор терминации облегчает отделение первичного транскрипта (пре-мРНК), комплементарного матрице, и РНК-полимеразы от матрицы. РНК-полимераза может вступить в следующий цикл транскрипции после присоединения субъединицы σ.
Ковалентная модификация (процессинг) матричной РНК.
Первичные транскрипты мРНК, прежде чем будут использованы в ходе синтеза белка, подвергаются ряду ковалентных модификаций. Эти модификации необходимы для функционирования мРНК в качестве матрицы.
Модификация 5'-конца.
К 5′-концу РНК добавляется кэп (метилированный гуаниновый нуклеотид), защищающий транскрипт от деградации.








