 2015-05-25
2015-05-25 669
669Спросите любого специализирующегося в нейронауках выпускника университета, какой организм больше всего использовался в клеточной биологии научения, и вы скорее всего услышите в ответ: аплизия. А на вопрос об исследователе, сыгравшем ключевую роль в этой программе работ, вам почти наверняка назовут имя Эрика Кэндела, профессора Фонда Говарда Хьюза в Нью-Йоркском колледже терапевтов и хирургов, автора одного из самых известных в последнем десятилетии пособий по нейронаукам [3] и плодовитого соавтора других книг, блестящего и неутомимого пропагандиста аплизии как идеального объекта для изучения памяти и редукционизма как методологической и философской концепции ее познания (Кэндел был столь ярым приверженцем редукционизма как философии и одновременно методологии, что однажды выступил перед аудиторией психблогов с лекцией на тему «Психотерапия и отдельный синапс» [4]).
Кэндел, получивший образование как психиатр, в шестидесятые годы какое-то время работал с аплизиями в Париже вместе с Ладиславом Тауком и имел возможность оценить перспективность этого моллюска при изучении вначале таких кратковременных процессов, как привыкание. В последующую четверть века этот организм стал главным объектом его самостоятельных исследований в Нью-Йорке, где в возглавляемой им лаборатории Колумбийского университета выросло несколько поколений специалистов-нейробиологов. Несомненно, Кэндел и его школа внесли важный вклад в нейробиологию кратковременных проявлений памяти как в экспериментальном плане, так и в смысле повышения престижа того раздела физиологии, к которому после его упадка в шестияесятые годы многие относились весьма сдержанно. Далеко не просто складывались личные отношения между учеными, работавшими с аплизией, и между всей этой группой и другими группами,, которые использовали иные модели, в частности с Дэном Элконом и его коллегами по работе с Hermissenda в Вудс-Хоуле (см. главу 7). Одно время трения стали столь заметными, что побудили автора научно-популярных работ Сюзан Олпорт посвятить им отдельную книгу [5]. Тем не менее общее направление исследований Кэндела и теоретические предпосылки, разработанные им в 70-х и 80-х годах, до недавнего времени не встречали особых возражений. Однако результаты, полученные в последние несколько лет в его собственной и в других лабораториях, побуждают расширить прежний несколько упрощенный редукционистский подход к интерпретации получаемых данных. Чтобы дать представление о существе этой критики, мне придется сначала рассмотреть редукционистскую концепцию Кэндела в ее наиболее выраженной форме.
В главе 7 уже говорилось, почему исследования на аплизии приобрели стратегическое значение для нейробиологии некоторых важных форм памяти. У этого животного очень несложный поведенческий репертуар, включающий различные виды научения, а их центральную нервную систему сравнительно нетрудно картировать, так как она состоит не более чем из 20 000 нейронов, сгруппированных в ряд обособленных ганглиев, и среди этих нейронов есть очень крупные клетки, легко распознаваемые индивидуально у всех особей. Ключевым моментом в подходе Кэндела было изучение простейших поведенческих реакций у интактного животного. Речь идет об элементарном рефлексе - втягивании жабры и сифона - с явлениями привыкания и сенситизации. Эту форму поведения или ее нервный механизм можно «отделить» от других поведенческих реакций путем последовательного уменьшения группы изучаемых в эксперименте нейронов. Предельным случаем будет наблюдение над двумя определенными нейронами, которые можно отпрепарировать, а затем вызвать образование синаптической связи в лабораторной чашке. По мнению Кэндела, взаимодействие таких нейронов и их реакции на добавляемые извне нейромедиаторы можно считать моделью памяти для рефлекса в ее крайне редуцированной форме. Каковы же доводы в пользу этого?
Рассмотрим сначала сам рефлекс. Аплизия дышит с помощью жабры, которая размещается в полости на верхней (дорсальной) стороне тела. В задней части этой полости имеется вырост в виде трубки - сифон. Если прикоснуться к области вокруг жабры или сифона, они втягиваются в полость: это простой защитный рефлекс. Нервный механизм такого рефлекса включает небольшое число (около 50) сенсорных нейронов, реагирующих на тактильное раздражение упомянутой области. Эти сенсорные нейроны контактируют примерно с 20 моторными нейронами как непосредственно, так и через промежуточные нейроны (интернейроны). Моторные нейроны расположены в абдоминальном (брюшном) ганглии и в свою очередь образуют синапсы на мышечных волокнах, втягивающих сифон и жабру. Эта относительно простая цепь схематически показана на рис. 9.2. Однократное раздражение поверхности тела около жабры или сифона приводит к их втягиванию, а при многократной стимуляции наступает привыкание; реакция на повторные стимулы постепенно ослабевает вплоть до ее полного исчезновения на некоторое время. Привыкание можно снять (дегабитуация, или сенситизация) сильным раздражением другого участка тела, например «хвоста», что приводит к полному восстановлению первоначальной реакции. Такие изменения рефлекса носят кратковременный и довольно неспецифический характер, и их следует рассматривать как формы неассоциативного научения. Наряду с этим, как показал Кэндел, возможна и выработка классического условного рефлекса, когда безусловным стимулом служит резкое воздействие на хвостовую область, а условным - слабое тактильное раздражение сифона. Слабая стимуляция обычно приводит и к слабо выраженному втягиванию сифона, тогда как после закрепления условного рефлекса такое же слабое раздражение вызывает сильную реакцию. Этот эффект сохраняется довольно долго, и поскольку здесь существует специфическая связь между стимулом и реакцией, его можно считать формой истинного ассоциативного научения.
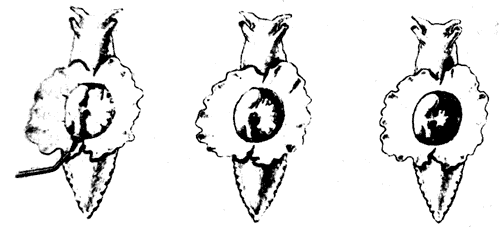
Рис. 9.2а. Рефлекс втягивания жабры и сифона у аплизии. Показано интактное животное. На сифон через трубку направлена струя воды (слева), это приводит к втягиванию сифона и фабры (в середине и справа).
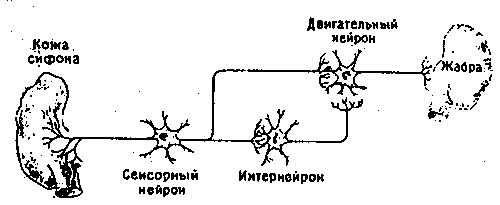
Рис. 9.2b. Схема нервных связей в «редуцированном» препарате.
Кэндел и его сотрудники занялись поисками нервных путей, участвующих в реакции втягивания жабры и сифона. Они использовали простые классические методы нейрофизиологии, а дальнейшая стратегия состояла в последовательном уменьшении числа нейронов. Для более точного контроля и количественной оценки реакции моллюсков обездвиживали, прикрепляя к пластинке, а для стандартизации тактильных стимулов применяли струю воды, направляемую через остроконечную трубку. Сокращения жабры можно было также прямо регистрировать с помощью фотоэлемента. Выявив нервную сеть, которая обеспечивала рефлекторную реакцию, исследователи смогли перейти ко второму вопросу: какая часть этой сети ответственна за привыкание? Не происходят ли при этом изменения в каких-то определенных клетках или синапсах? Этот вопрос, очевидно, имеет отношение к описанному выше первому критерию. Но, поскольку группа Кэндела состояла из нейрофизиологов, а не биохимиков, исследователи начали с изучения электрических свойств клеток, что соответствует моему шестому критерию.
Однако для получения ответа требовалось еще одно упрощение изучаемой системы, в результате которого живые аплизии превращались в неактивные, удобные для манипуляций «препараты». Животное можно вскрыть, обнажив абдоминальный ганглий, или даже полностью изолировать этот ганглий вместе с нервами, участком кожи и жаброй. Проведя такую операцию, Кэндел мог считать, что исключил все посторонние источники сигналов - другие периферические нервы, нейро-модуляторы, циркулирующие в крови и т. п. При этом он получил возможность выявлять крупные клеточные тела моторных нейронов и, как говорилось в главе 7, повторно идентифицировать «те же самые» клетки при смене одного животного другим (рис. 9.2). В результате операции живой моллюск становился чем-то вроде компьютерной схемы, и ученые могли исследовать ее свойства как электротехники, получившие новое оборудование и пытающиеся понять принципы его работы. В такой системе осязательный стимул для поведенческих реакций можно заменить его нейрофизиологическим аналогом, т. е. прямым электрическим раздражением нервов, идущих к сенсорным нейронам. Точно так же и мышечную реакцию - втягивание жабры и сифона - можно вызывать прямой стимуляцией нервов, отходящих от моторных нейронов к жаберной мускулатуре.
На следующем этапе работы изолированные и «упрощенные» препараты использовались для того, чтобы выяснить локализацию механизмов привыкания, т. е. найти те звенья нервной цепи, которые начинают слабее отвечать на повторное раздражение. К середине семидесятых годов стало ясно, что ни подходящие к сенсорным нейронам, ни отходящие от двигательных нейронов нервы не обладают свойствами, позволяющими объяснить привыкание, так как их электрические ответы не уменьшаются. Следовательно, клетки, ответственные за привыкание, должны занимать промежуточное положение в сенсорно-моторной цепи абдоминального ганглия. И действительно, регистрация электрических сигналов от моторных нейронов в этом ганглии в период привыкания выявила постепенное уменьшение их частоты. Из этого был сделан вывод, что «место» привыкания лежит между сенсорными и моторными нейронами [6].
Однако даже в таком крайне упрощенном препарате имеется множество нервных цепей и многие тысячи клеток. Сенсорные нейроны, например, имеют прямые и опосредованные связи с моторными нейронами. Прямые связи обеспечиваются синапсами между аксоном сенсорного нейрона и дендритом или телом моторного нейрона (моносинаптический путь); в непрямом, полисинаптическом пути сначала сенсорный нейрон устанавливает синаптическую связь с интернейроном, а тот в свою очередь образует синапс с моторным нейроном. (Здесь следует ввести еще два соотносящиеся друг с другом термина. Когда одна клетка воздействует на другую непосредственно путем модификации синапса, образуемого ею на второй клетке, говорят о гомосинаптическом эффекте; если же действие одной клетки на другую модулируется третьей клеткой, имеющей синапсы с двумя первыми, такой эффект называют гетеросинаптическим.)
Анализ активности моторных нейронов аплизии после сенсорной стимуляции показывает, что они реагируют как непосредственно, т. е. моносинаптически, так и полисинаптически, через посредство интернейронов. На просьбу указать наиболее вероятное место синаптической пластичности теоретики скорее всего выберут интернейроны, поскольку они, очевидно, могут получать и модулировать сигналы, поступающие по множеству различных входов, прежде чем направить их по различным выходным путям. Простая модель научения, предложенная Хеббом и представленная на рис. 6.1, требует участия трех нейронов. Иными словами, если она верна, то предполагает гетеросинаптическое взаимодействие. Однако, к удивлению многих, в начале восьмидесятых годов группа Кэндела показала, что элемент, ответственный за привыкание, чрезвычайно прост: это прямая синаптическая связь между сенсорным и моторным нейронами; в частном случае - синапс между сенсорным нейроном и одним определенным очень крупным моторным нейроном, причем связь здесь моносинаптическая с модуляцией гомосинаптического типа.
Постепенное упрощение препарата до изолированной нервной цепи подготовило почву для его предельной редукции: сотрудник Кэндела Самюэл Шахер выделил два специфических нейрона - сенсорный и двигательный - и поместил их вместе в чашку Петри, т. е. перенес в условия тканевой культуры. Уже давно известно, что в этих условиях нейроны, подобно другим клеткам, много дней и даже недель остаются живыми, если поддерживать нужную температуру, аэрировать среду и снабжать клетки глюкозой и другими необходимыми веществами. При этом многие виды культивируемых клеток продолжают делиться, а нейроны хотя и не делятся, но растут и образуют аксоны, дендриты и даже синаптические соединения. В культурах Шахера сенсорные нейроны формировали синапсы на моторных нейронах, и их электростимуляция вызывала электрическую реакцию последних. При многократном раздражении сенсорной клетки ответ моторного нейрона постепенно затухал, т. е. развивалось привыкание [7]. В конце концов в лаборатории Кэндела было получено то, что он описал как изолированный синапс, «обучающийся в блюдце». Фактически это был полный триумф редукционистской стратегии, которой следовала группа Кэндела. Это, казалось, оправдывало его претензию на постановку такой цели, как расшифровка «клеточного алфавита» научения. Сенсорно-моторный синапс, безусловно, может считаться одной из букв такого алфавита.
Все, о чем шла речь до сих пор, относится в основном к области нейрофизиологии. А что можно сказать о биохимических механизмах реакций на изученных Кэнделом уровнях клеточной организации? Если привыкание происходит вследствие ослабления постсинаптической реакции в отдельном синапсе, то по логике вещей оно должно быть результатом либо пресинаптического, либо постсинаптического процесса или, разумеется, их сочетания. Например, может постепенно уменьшаться количество нейромедиатора, выделяемое пресинаптической клеткой, или происходить изменение рецепторов на постсинаптической стороне, так что они будут слабее реагировать на прежние дозы медиатора; возможно и одновременное участие обоих механизмов. Вопрос об относительной роли пре- и постсинаптической пластичности в последние годы стал одним из важных предметов спора, но большинство теоретиков склонно было приписывать пластичность в основном постсинаптической стороне.
Первой задачей нейрохимического исследования для группы, работавшей с аплизией, стала идентификация медиатора, передающего сигналы в синапсе. Им оказалось широко распространенное вещество серотонин, или 5-гидрокситриптамин. К середине семидесятых годов сотрудники группы установили, что в период привыкания в изолированном ганглии происходит неуклонное снижение секреции серотонина пресинаптическими окончаниями сенсорных клеток, тогда как чувствительность постсинаптических рецепторов этого вещества не изменяется. Пониженная секреция серотонина сопровождалась также изменением свойств пресинаптической мембраны, в особенности уменьшением трансмембранного переноса кальция внутрь клетки (биохимическое значение этого станет понятным позже). В параллельных экспериментах было показано, что сенситизация (т. е. эффект, в известном смысле противоположный привыканию; см. главы 6 и 7) тоже связана с пресинаптическими процессами, но здесь уже необходимо усиление секреции серотонина и притока кальция в клетки. Тот факт, что оба процесса - и привыкание, и сенситизация - происходят с участием пресинаптических механизмов, оказался довольно неожиданным для тех, кто моделирует нервные функции.
Десятилетие спустя были проведены аналогичные эксперименты на клетках в культуре, и они дали те же результаты. На этот раз для «обучения» не требовалось даже двух клеток, реакция полностью воспроизводилась при воздействии серотонином на изолированный моторный нейрон. Дальнейшее упрощение трудно придумать!
В следующей главе я гораздо подробнее остановлюсь на биохимии этих процессов в контексте моих собственных экспериментов. Сейчас я не буду вдаваться в детали, а хочу лишь подчеркнуть то, что Кэндел объясняет рефлекс, привыкание и сенситизацию, переходя к все более простым системам. Сначала он сводит сложную форму поведения интактного организма - втягивание сифона и жабры - к реакции нервной цепи, в которой в результате взаимодействия всего лишь двух клеток развивается привыкание, а потом интерпретирует электрическую (т. е. физиологическую) реакцию синапсов как каскад биохимических процессов в пресинаптическом нейроне.








