 2015-06-14
2015-06-14 576
576В предыдущих подразделах рассматривались функции отдельных клеток, направленные на противодействие осмотическому и токсическому эффектам солей. В этом подразделе показано, как защитные механизмы отдельных клеток складываются в единую систему, позволяющую решать эти задачи в стрессовых условиях на уровне целого организма. Как указывалось выше, одна из основных стратегий в адаптации к высоким концентрациям солей, проявляющаяся на клеточном уровне, состоит в поддержании низких концентраций Na+ и С1 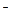 в цитоплазме и создании таким путем благоприятного микроокружения для функционирования цитоплазматических биополимеров. Одновременно за счет биосинтеза осмолитов в цитоплазме и депонирования этих ионов в вакуолях клетки поддерживают ОСВ на необходимом для жизнеобеспечения уровне.
в цитоплазме и создании таким путем благоприятного микроокружения для функционирования цитоплазматических биополимеров. Одновременно за счет биосинтеза осмолитов в цитоплазме и депонирования этих ионов в вакуолях клетки поддерживают ОСВ на необходимом для жизнеобеспечения уровне.
С появлением в ходе эволюции многоклеточных организмов, дифференцированных на органы и ткани, наряду с прежней появилась новая стратегия в водном обмене и ионном гомеостатировании. Стало важным, во-первых, поддерживать водный ток в системе: почва —корень—побег, во-вторых, избегать накопления ионов, присутствующих в среде в высоких концентрациях, в молодых, активно метаболизирующих частях растения, прежде всего в меристе-матических тканях и генеративных органах. Известно, что в меристематических клетках, цветках, семенах и плодах содержание Na+ и С1 в расчете на сырую биомассу не превышает нескольких ммоль/кг даже при сильном почвенном засолении.
|
|
|
Вода и растворенные в ней вещества движутся из почвенного раствора в клетки эпидермиса и коры. Далее они транспортируются в радиальном направлении к центру корня и затем по ксилеме вверх, в листья. Движение ионов в радиальном направлении корня, так же как и движение воды (см. гл. 5), может осуществляться по апопласту (апопластный путь), и через клетки (путь от клетки к клетке). За счет функционирования локализованных в ПМ и тонопласте ион-транспортирующих белков формируется ионный состав цитоплазмы клеток корня, в котором содержание Na+ и С1 ограничено, а К+ доминирует над Na+. Значительная часть ионов, передвигающаяся в радиальном направлении корня, достигнув эндодермы, вынуждена из-за поясков Каспари пересекать ПМ и входить в цитоплазму клеток этой ткани, однако наличие обходных путей (пропускные клетки эндодермы и незрелые ткани кончика корня) позволяет ионам передвигаться и через апопласт. Эндодерма, таким образом, не может быть абсолютным барьером для ионов, хотя мембраны ее клеток вносят существенный вклад в регуляцию ионных потоков через корень. В связи с этим, когда интенсивность транспирации высока, апопластный путь играет заметную роль в транспорте Na+ и С1 . Ионы движутся в этом случае с массовым потоком
|
|
|
воды и растворенных веществ по градиенту гидростатического давления (гидравлический поток). При низкой интенсивности транспирации движение Na+ и С1 осуществляется преимущественно от клетки к клетке. В этот путь движения ионов вовлечены ионные каналы, переносчики и ион-транспортирующие АТФазы. Для транспорта ионов в растении очень важен этап загрузки ионами ксилемы. Функцию загрузки выполняет плазматическая мембрана паренхимных клеток, окружающих ксилему. Показано, что у гликофитов в норме основным по массе катионом, транспортируемым в ксилему, является К+, а перенос К+ из клеток обкладки в ксилему осуществляется через активируемые при деполяризации ПМ выходные К+-каналы. У галофитов, а также при высокой солености среды и у гликофитов из транспортируемых в ксилему катионов доминирует Na+. Установлено, что у A. thaliana перенос ионов Na+ из паренхимных клеток обкладки в ксилему осуществляет Nа+/Н+-антипортер ПМ этих клеток, кодируемый геном SOS1. У ячменя в паренхимных клетках, прилегающих к сосудам ксилемы корня, идентифицированы анионные каналы, через которые происходит загрузка ксилемы ионами С1 .Исследование транспорта С1 через эти каналы показало, что они относятся к анионным каналам так называемого «медленного» типа. Кинетика транспорта С1 через них характеризуется секундным диапазоном.
При движении ксилемного сока вверх между ним и клетками, прилегающими к ксилеме, происходит обмен ионов, в результате которого ионный состав раствора, достигающего клеток листьев, существенно отличается от ионного состава первичного ксилемного сока в корне. Оказавшись в листьях, ионы Na+ и С1 проникают в клетки и там депонируются в вакуолях, повышая осмотическое давление вакуолярного сока. Процесс депонирования Na+ и С1 в вакуолях наиболее ярко выражен у галофитов. Следует отметить, что депонирование Na+ и С1 наблюдается также и у гликофитов, которые характеризуются относительно высокой устойчивостью к водному дефициту и высоким концентрациям солей.
Согласованное функционирование ион-транспортирующих систем в отдельных клетках обеспечивает при стрессах поддержание низких концентраций Na+ и С1 в цитоплазме самих клеток, низкое содержание Na+ и С1 в генеративных органах и активно метаболизирующих тканях и вносит вклад в формирование градиентов водного потенциала в системе: почва — корень — побег, путем накопления Na+ и С1 в вакуолях.
Как уже отмечалось, стратегия ионного транспорта и осморегуляции, основанная на накоплении ионов в вакуолях надземных органов, характерна для большинства галофитов, в том числе и тех, чьи листья снабжены солевыводящими, как у кермека (Limonium gmelinii), и солелокализующими, как у М. crystallinum, железами. Соль в вакуолях, занимающих большую часть клеточного объема, является «дешевым» осмотиком, тогда как «дорогие» осмолиты органической природы поддерживают осмотическое давление в цитоплазме. В связи с функцией накопления ионов в вакуолях важное значение у галофитов имеет баланс потоков Na+ и С1 из почвенного раствора в корни, с одной стороны, и из корней в побеги, с другой. Количество ионов, переносимое в побеги, должно быть достаточным для поддержания осмотического давления в вакуолях на нужном уровне. Если скорость аккумуляции ионов в вакуолях клеток надземных органов галофитов высока, т. е. высока активность Na+/Н+-антипортера тонопласта, доставка ионов из корней в побеги в достаточных количествах приводит к стимуляции их роста.
Гликофиты в условиях почвенного засоления не способны поддерживать низкие концентрации Na+ и С1 в цитоплазме и аккумулировать ионы в вакуолях своих клеток столь эффективно, как это делают галофиты. Большое значение в формировании признака солеустойчивости у гликофитов (в целом более низкой, чем у галофитов) имеет ограничение входящих потоков Na+ и С1 в корни. При этом осморегуляция их клеток, даже в вакуолярном компартменте, осуществляется в значительной степени за счет осмолитов органической природы. Однако способность гликофитов синтезировать осмолиты весьма ограничена. Распределение поступающих из почвенного раствора ионов между корневой системой и надземными органами у гликофитов определяется, как и у галофитов, функцией паренхимных клеток обкладки ксилемы корня, которые могут ограничивать поступление ионов в надземные органы. Различные виды и сорта растений различаются по эффективности этой функции. У сои (Glycine max) и пшеницы (Triticum aestivum) различия между сортами в ограничении ионных потоков из корней в побеги определяются одним геном. Генетический анализ показал, что такой ген KNA1 у пшеницы локализован в хромосоме 4D.
|
|
|
Согласованное действие ион-транспортирующих белков в мембранах клеток различных органов и тканей, поддерживающих ионный гомеостаз и формирующих ионные градиенты в системе целого растения, подразумевают орган- и тканcспецифическую экспрессию ион-транспортирующих белков. Биосинтез осмолитов в растении также регулируется органо- и тканcспецифическим образом. Тот или иной белок, индуцируемый стрессом, должен появиться не только в нужном месте, но и в нужное время. Важную роль в пространственной и временной регуляции экспрессии стрессовых генов играют гормоны и орган-и тканеспецифические промоторы.
К механизмам адаптации к засолению и водному дефициту, которые проявляются на уровне целого растения, относятся С4- и САМ-пути фиксации углерода (см. гл. 3). Транспирационный ток воды многократно ускоряет поступление в растения ионов Na+ и С1 , в избытке содержащихся в почвенном растворе при засолении. Полностью открытые устьица, поддерживающие высокую интенсивность транспирации, создавали бы не только угрозу потери воды растением, но и ускоряли бы накопление Na+ и С1 в цитоплазме клеток. При высоких концентрациях соли в среде и водном дефиците апертура устьичных щелей, однако, уменьшается, что делает СО2 менее доступным для растений и приводит к подавлению фотосинтеза. В связи с этим у многих представителей флоры в зоне аридного климата, обитающих при недостатке влаги и избытке солей в почве, фотосинтез идет по С4- или САМ-пути. Преимущество С4- и САМ-метаболизма в условиях водного дефицита и почвенного засоления состоит в возможности осуществлять фотосинтез при высоком устьичном сопротивлении. Можно считать поэтому, что С4- и САМ-пути — это такие способы фотосинтеза, которые не только экономят воду при низком водном потенциале почвенного раствора, но и препятствуют избыточному поглощению растениями Na+ и С1 . У некоторых растений, в частности у галофита М. cristallinum, высокие концентрации NaCl в почве приводят к индукции ферментов САМ-пути.
|
|
|








