 2015-06-16
2015-06-16 11274
11274К цианобактериям относится большая группа организмов, сочетающих прокариотное строение клетки со способностью осуществлять фотосинтез, сопровождающийся выделением O2, что свойственно разным группам водорослей и высших растений. Объединение черт, присущих организмам, относящимся к разным царствам или даже надцарствам живой природы, сделало цианобактерии объектом борьбы за принадлежность к низшим растениям (водорослям) или бактериям (прокариотам).
Вопрос о положении цианобактерии (сине-зеленых водорослей) в системе живого мира имеет долгую и противоречивую историю. В течение длительного времени они рассматривались как одна из групп низших растений, поэтому и систематика осуществлялась в соответствии с правилами Международного кодекса ботанической номенклатуры. И только в 60-х гг. XX в., когда было установлено четкое различие между прокариотным и эукариотным типами клеточной организации и на основании этого К. ван Нилем и Р. Стейниером сформулировано определение бактерий как организмов, имеющих прокариотное строение клетки, встал вопрос о пересмотре положения сине-зеленых водорослей в системе живых организмов.
Изучение цитологии клеток сине-зеленых водорослей с помощью современных методов привело к неоспоримому выводу о том, что эти организмы также являются типичными прокариотами. Как следствие этого Р. Стейниером было предложено отказаться от названия "сине-зеленые водоросли" и называть данные организмы "цианобактериями" — термином, отражающим их истинную биологическую природу. Воссоединение цианобактерий с остальными прокариотами поставило исследователей перед необходимостью пересмотра существующей классификации этих организмов и подчинения ее правилам Международного кодекса номенклатуры бактерий.
В течение длительного времени альгологами было описано около 170 родов и больше 1000 видов сине-зеленых водорослей. В настоящее время ведется работа по созданию новой систематики цианобактерий, основанной на изучении чистых культур. Уже получено больше 300 чистых штаммов цианобактерий. Для классификации использованы постоянные морфологические признаки, закономерности развития культуры, особенности клеточной ультраструктуры, величина и нуклеотидная характеристика генома, особенности углеродного и азотного метаболизма и ряд других.
Цианобактерий — морфологически разнообразная группа грамотрицательных эубактерий, включающая одноклеточные, колониальные и многоклеточные формы. У последних единицей структуры служит нить (трихом, или филамент). Нити бывают простые или ветвящиеся. Простые нити состоят из одного ряда клеток (однорядные трихомы), имеющих одинаковые размеры, форму и строение, или клеток, различающихся по этим параметрам. Ветвящиеся трихомы возникают в результате разных причин, в связи с чем различают ложное и истинное ветвление. К истинному ветвлению приводит способность клеток трихома делиться в разных плоскостях, в результате чего возникают многорядные трихомы или однорядные нити с однорядными же боковыми ветвями. Ложное ветвление трихомов не связано с особенностями деления клеток внутри нити, а есть результат прикрепления или соединения разных нитей под углом друг к другу.
В процессе жизненного цикла некоторые цианобактерий формируют дифференцированные единичные клетки или короткие нити, служащие для размножения (баеоциты, гормогонии), выживания в неблагоприятных условиях (споры, или акинеты) или азотфиксации в аэробных условиях (гетероцисты). Более подробная характеристика дифференцированных форм цианобактерий дана ниже при описании их систематики и процесса азотфиксации. Краткая характеристика акинет представлена в гл. 5. Для разных представителей этой группы характерна способность к скользящему движению. Оно свойственно как нитчатым формам (трихомы и/или гормогонии), так и одноклеточным (баеоциты).
Известны разные способы размножения цианобактерий. Деление клеток происходит путем равновеликого бинарного деления, сопровождающегося образованием поперечной перегородки или перетяжки; неравновеликого бинарного деления (почкования); множественного деления (см. рис. 20, А–Г). Бинарное деление может происходить только в одной плоскости, что у одноклеточных форм приводит к образованию цепочки клеток, а у нитчатых — к удлинению однорядного трихома. Деление в нескольких плоскостях ведет у одноклеточных цианобактерий к формированию скоплений правильной или неправильной формы, а у нитчатых — к возникновению многорядного трихома (если к такому делению способны почти все вегетативные клетки нити) или однорядного трихома с боковыми однорядными ветвями (если способность к делению в разных плоскостях обнаруживают только отдельные клетки нити). Размножение нитчатых форм осуществляется также с помощью обрывков трихома, состоящих из одной или нескольких клеток, у некоторых — также гормогониями, отличающимися по ряду признаков от трихомов, и в результате прорастания акинет в благоприятных условиях.
Начатая работа по классификации цианобактерий в соответствии с правилами Международного кодекса номенклатуры бактерий привела к выделению 5 основных таксономических групп в ранге порядков, различающихся морфологическими признаками (табл. 27). Для характеристики выделенных родов привлечены также данные, полученные при изучении клеточной ультраструктуры, генетического материала, физиолого-биохимических свойств.
К порядку Chroococcales отнесены одноклеточные цианобактерий, существующие в виде одиночных клеток или формирующие колонии (рис. 80). Для большинства представителей этой группы характерно образование чехлов, окружающих каждую клетку и, кроме того, удерживающих вместе группы клеток, т. е. участвующих в формировании колоний. Цианобактерий, клетки которых не образуют чехлов, легко распадаются до одиночных клеток. Размножение осуществляется бинарным делением в одной или нескольких плоскостях, а также почкованием.
Таблица 27. Основные таксономические группы цианобактерий
| Одноклеточные формы: одиночные клетки или колонии | Многоклеточные формы: нитчатые | |||
| Пор. Chroococcales | Пор. Pleurocapsales | Пор. Oscillatoriales | Пор. Nostocales | Пор. Stigoneomatales |
| Размножение бинарным делением в одной или более плоскостях или почкованием | Размножение множественным делением или чередованием бинарного и множественного деления | Трихомы неветвящиеся, состоят из одного ряда только вегетативных клеток. Рост трихома — делением клеток в одной плоскости | В неветвящихся однорядных трихомах помимо вегетативных образуются дифференцированные клетки: гетероцисты и иногда акинеты. Рост трихома — делением клеток в одной плоскости | Те же признаки, что и у представителей пор. Nostocales. Отличительный признак: способность вегетативных клеток трихома к делению более чем в одной плоскости, приводящему к появлению многорядных трихомов или трихомов с истинным ветвлением |
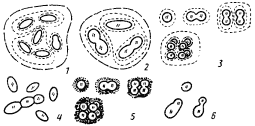 |
| Рис. 80. Цианобактерии, отнесенные к порядку Chroococcales: 1 — Gloeobacter; 2 — Gloeothece; 3 — Gloeocapsa; 4 — Synechococcus; 5 — Synechocystis; 6 — Chamaesiphon. Прерывистой линией обозначены чехлы, точками — капсулы, черточками — тилакоиды |
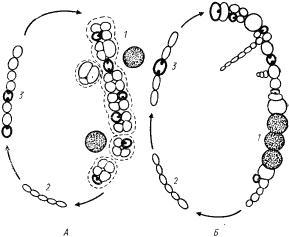
В порядок Pleurocapsales выделены одноклеточные цианобактерий, способные к размножению путем множественного деления. Они существуют в виде одиночных клеток или скоплений, удерживаемых вместе с помощью внешнего (по отношению к наружной мембране) фибриллярного слоя клеточной стенки. Скопления могут состоять всего из нескольких клеток разной формы, иметь кубическую или неправильную форму. Входящие в эту группу цианобактерий различаются способностью размножаться только множественным делением или чередованием процессов бинарного и множественного деления (рис. 81). Освобождающиеся в результате множественного деления баеоциты могут быть подвижными или неподвижными. У подвижных баеоцитов при освобождении из материнской клетки отсутствует дополнительный фибриллярный слой клеточной стенки. Подвижность их теряется, когда этот слой синтезируется. У неподвижных форм к моменту выхода из материнской клетки дополнительный слой клеточной стенки уже сформирован.
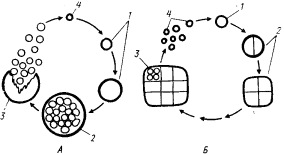 |
| Рис. 81. Циклы развития некоторых цианобактерий, включенных в порядок Pleurocapsales. А — цикл развития цианобактерий рода Dermocarpa: 1 — увеличение объема баеоцита до размеров вегетативной клетки; 2 — множественное деление, приводящее к образованию баеоцитов; 3 — разрыв материнской клетки и освобождение подвижных баеоцитов; 4 — синтез внешнего слоя клеточной стенки и потеря баеоцитом подвижности. Б — цикл развития цианобактерий рода Chroococcidiopsis: I — увеличение объема баеоцита до размеров вегетативной клетки; 2 — серия бинарных делений больше чем в одной плоскости; 3 — множественное деление части клеток колонии; 4 — освобожденные неподвижные баеоциты. Подвижные баеоциты обведены сплошной тонкой линией; неподвижные — сплошной жирной линией. Объяснения см. в тексте |
В отличие от рассмотренных выше цианобактерий в последующие порядки включены многоклеточные формы, имеющие нитчатое строение. Особенностью цианобактерий, отнесенных в порядок Oscillatoriales, является недифференцированность трихома (последний состоит только из вегетативных клеток) и его рост путем деления клеток в одной плоскости. Цианобактерии этой таксономической группы различаются строением трихомов и отдельных клеток, особенностями соединения клеток в трихоме, наличием или отсутствием чехла, способностью к движению и некоторыми другими морфологическими признаками (рис. 82). Для большинства представителей этой группы характерны прямые трихомы, клетки в которых дисковидные или цилиндрические плотно прилегают друг к другу или разделены глубокой перетяжкой. Трихомы могут быть окружены общим чехлом разной толщины. Скользящее движение свойственно цианобактериям, не образующим чехла или со слабым развитием последнего. К этой же группе относятся цианобактерии, имеющие подвижные спиралевидные трихомы, состоящие из клеток разной формы, не окруженные чехлом.
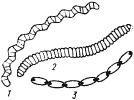 |
| Рис. 82. Цианобактерии, включенные в порядок Oscillatoriales: 1 — Spirulina; 2 — Oscillatoria; 3 — Pseudoanabaena |
Дальнейший шаг по пути морфологического усложнения сделан цианобактериями, включенными в порядок Nostocales (рис. 83). Они представлены однорядными неветвящимися нитями, рост которых происходит путем деления клеток в одной плоскости (под прямым углом к длинной оси трихома). При культивировании на среде без связанного азота некоторые вегетативные клетки дифференцируются в гетероцисты — центры азотфиксации в аэробных условиях. Ряд представителей группы образует акинеты — единственный тип покоящихся форм у цианобактерий. Размножение происходит короткими обрывками трихомов, морфологически не отличающимися от зрелых длинных нитей, и в результате прорастания акинет, если последние образуются. У некоторых цианобактерий в дополнение к описанным выше способам размножения для этой цели служат гормогоний. Последние представляют собой короткие нити, отличающиеся рядом морфологических признаков от родительского трихома: они состоят из небольшого числа мелких активно движущихся вегетативных клеток, иногда иной формы, чем клетки родительского трихома; могут содержать газовые вакуоли; никогда не окружены чехлом. Основное отличие гормогониев от зрелых и молодых трихомов — отсутствие гетероцист, даже если культура находится в среде, не содержащей связанного азота. Для выделения отдельных родов использованы такие признаки, как расположение гетероцист и акинет в нити, форма вегетативных клеток. В частности, трихом цианобактерий рода Calothrix образован клетками разной ширины, т. е. имеет асимметричное строение.
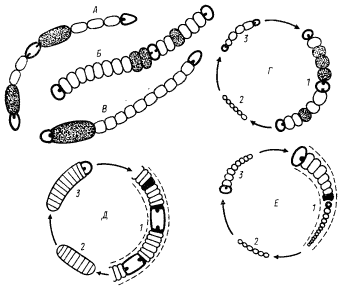 |
| Рис. 83. Цианобактерии, отнесенные к порядку Nostocales: А — Anabaena; Б — Nodularia; В — Cylindrospermum; Г — Nostoc; Д, — Scytonema; Е — Calothrix: 1 — трихом в зрелом состоянии; 2 — гормогоний; 3 — молодой трихом. Гетероцисты изображены в виде клеток с толстой клеточной стенкой и полярными гранулами; акинеты — в виде темных клеток. Прерывистой линией вдоль трихома обозначен чехол |
В порядок Stigonematales объединены цианобактерий, отличающиеся от представителей предыдущего порядка способностью вегетативных клеток трихома к делению более чем в одной плоскости (рис. 84). К этой группе отнесено несколько родов, различающихся циклами развития, расположением гетероцист в трихомах и некоторыми другими признаками.
Электронно-микроскопическое изучение вегетативных клеток цианобактерий обнаружило принципиальное сходство их строения с клетками грамотрицательных эубактерий. Более чем у 200 чистых культур определен состав оснований хромосомной ДНК. По этому признаку цианобактерии обнаруживают гетерогенность (молярное содержание ГЦ-оснований в ДНК от 35 до 71%), сравнимую только с остальными прокариотами (25-75%).
 |
| Рис. 84. Цианобактерии порядка Stigonematales: А — Chlorogloeopsis; Б — Fischerella: 1 — зрелый трихом; 2 — гормогоний; 3 — молодой трихом. Обозначение гетероцист, акинет и чехла см. на рис. 83 |
В качестве одной из примечательных особенностей генетического материала цианобактерии отмечают значительные различия величины цианобактериальной хромосомы. Размеры геномов, изученные более чем у 100 штаммов из разных групп, располагаются в диапазоне 1,6 — 8,6-109 Да, при этом просматривается определенная корреляция между степенью морфологической сложности и величиной генома, достигающего максимальных значений у цианобактерии со сложной организацией трихомов и циклами развития. В группе цианобактерии сформирован самый крупный геном, обнаруженный до сих пор у прокариот. В то же время некоторые цианобактерии в отношении морфологической сложности также достигли вершины в мире прокариот и не имеют равных среди других грамотрицательных эубактерий.
Клетки цианобактерии, за исключением принадлежащих к роду Gloeobacter, характеризуются развитой системой внутрицитоплазматических мембран (тилакоидов), в которых локализованы компоненты фотосинтетического аппарата. Единственная энергопреобразующая мембрана Gloeobacter — цитоплазматическая, где локализованы процессы фотосинтеза и дыхания.
Большой интерес представляет группа цианобактерии из-за сосредоточения в ней разнообразных физиологических возможностей. В недрах этой группы, вероятно, сформировался и в целом оформился фотосинтез, основанный на функционировании двух фотосистем, характеризующийся использованием H2O в качестве экзогенного донора электронов и сопровождающийся выделением О2.
У цианобактерии обнаружена способность к бескислородному фотосинтезу, связанная с отключением II фотосистемы при сохранении активности I фотосистемы (см. рис. 75, В). В этих условиях у них возникает потребность в иных, чем H2O, экзогенных донорах электронов. В качестве последних цианобактерии могут использовать некоторые восстановленные соединения серы (H2S, Na2S2O3), H2, ряд органических соединений (сахара, кислоты). Так как поток электронов между двумя фотосистемами прерывается, синтез АТФ сопряжен только с циклическим электронным транспортом, связанным с I фотосистемой. Способность к бескислородному фотосинтезу обнаружена у многих цианобактерии из разных групп, но активность фиксации CO2 за счет этого процесса низка, составляя, как правило, несколько процентов от скорости ассимиляции CO2 в условиях функционирования обеих фотосистем. Только некоторые цианобактерии могут расти за счет бескислородного фотосинтеза, например Oscillatoria limnetica, выделенная из озера с высоким содержанием сероводорода. Способность цианобактерии переключаться при изменении условий с одного типа фотосинтеза на другой служит иллюстрацией гибкости их светового метаболизма, имеющей важное экологическое значение.
Хотя подавляющее большинство цианобактерии являются облигатными фототрофами, в природе они часто находятся длительное время в условиях темноты. В темноте у цианобактерии обнаружен активный эндогенный метаболизм, энергетическим субстратом которого служит запасенный на свету гликоген, катаболизируемый по окислительному пентозофосфатному циклу, обеспечивающему полное окисление молекулы глюкозы. На двух этапах этого пути с НАДФ-H2 водород поступает в дыхательную цепь, конечным акцептором электронов в которой служит O2.
O. limnetica, осуществляющая активный фотосинтез бескислородного типа, оказалась также способной в темноте в анаэробных условиях при наличии в среде серы осуществлять перенос электронов на молекулярную серу, восстанавливая ее до сульфида. Таким образом, анаэробное дыхание также может поставлять цианобактериям в темноте энергию. Однако насколько широко распространена такая способность среди цианобактерии, неизвестно. Не исключено, что она свойственна культурам, осуществляющим бескислородный фотосинтез.
Другой возможный путь получения цианобактериями в темноте энергии — гликолиз. У некоторых видов найдены все ферменты, необходимые для сбраживания глюкозы до молочной кислоты, однако образование последней, а также активности гликолитических ферментов низки. Кроме того, содержание АТФ в клетке в анаэробных условиях резко падает, так что, вероятно, жизнедеятельность цианобактерии только за счет субстратного фосфорилирования поддерживаться не может.
У всех изученных цианобактерии ЦТК из-за отсутствия -кетоглутаратдегидрогеназы "не замкнут" (рис. 85). В таком виде он не функционирует в качестве пути, ведущего к получению энергии, а выполняет только биосинтетические функции. Способность в той или иной степени использовать органические соединения для биосинтетических целей присуща всем цианобактериям, но только некоторые сахара могут обеспечивать синтез всех клеточных компонентов, являясь единственным или дополнительным к CO2 источником углерода.
Цианобактерии могут ассимилировать некоторые органические кислоты, в первую очередь ацетат и пируват, но всегда только в качестве дополнительного источника углерода. Метаболизирование их связано с функционированием "разорванного" ЦТК и приводит к включению в весьма ограниченное число клеточных компонентов (рис. 85). В соответствии с особенностями конструктивного метаболизма у цианобактерии отмечают способность к фотогетеротрофии или облигатную привязанность к фотоавтотрофии. В природных условиях часто цианобактерии осуществляют конструктивный метаболизм смешанного (миксотрофного) типа.
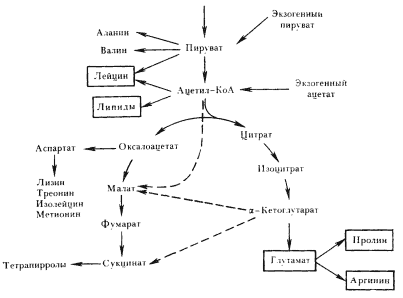 |
| Рис 85. "Разорванный" ЦТК у цианобактерий. Обведены продукты метаболизирования экзогенного ацетата. Пунктирными линиями изображены реакции глиоксилатного шунта |
Некоторые цианобактерии способны к хемогетеротрофному росту. Набор органических веществ, поддерживающих хемогетеротрофный рост, ограничен несколькими сахарами. Это связывают с функционированием у цианобактерий в качестве основного катаболического пути окислительного пентозофосфатного цикла, исходным субстратом которого служит глюкоза. Поэтому только последняя или сахара, ферментативно легко превращаемые в глюкозу, могут метаболизироваться по этому пути.
Одна из загадок метаболизма цианобактерий — неспособность большинства из них расти в темноте с использованием органических соединений. Невозможность роста за счет субстратов, метаболизируемых в ЦТК, связана с "разорванностью" этого цикла. Но основной путь катаболизма глюкозы — окислительный пентозофосфатный цикл — функционирует у всех изученных цианобактерий. В качестве причин называют неактивность систем транспорта экзогенных сахаров в клетку, а также низкую скорость синтеза АТФ, сопряженного с дыхательным электронным транспортом, вследствие чего количество вырабатываемой в темноте энергии достаточно только для поддержания клеточной жизнедеятельности, но не роста культуры.
Цианобактерии, в группе которых, вероятно, сформировался кислородный фотосинтез, впервые столкнулись с выделением O2 внутри клетки. Помимо создания разнообразных систем защиты от токсических форм кислорода, проявляющихся в устойчивости к высоким концентрациям O2, цианобактерии адаптировались к аэробному способу существования путем использования молекулярного кислорода для получения энергии.
В то же время для ряда цианобактерий показана способность расти на свету в строго анаэробных условиях. Это относится к видам, осуществляющим фотосинтез бескислородного типа, которых в соответствии с принятой классификацией следует отнести к факультативным анаэробам. (Фотосинтез любого типа по своей природе — анаэробный процесс. Это хорошо видно в случае фотосинтеза бескислородного типа и менее очевидно для кислородного фотосинтеза). Для некоторых цианобактерий показана принципиальная возможность протекания темновых анаэробных процессов (анаэробное дыхание, молочнокислое брожение), однако низкая активность ставит под сомнение их роль в энергетическом метаболизме цианобактерий. Зависимые и не зависимые от O2 способы получения энергии, обнаруженные в группе цианобактерий, суммированы в табл. 28.
Конструктивный метаболизм цианобактерий представляет собой шаг вперед по пути дальнейшей независимости от органических соединений внешней среды по сравнению с пурпурными и зелеными серобактериями. Для построения всех веществ клетки цианобактериям нужен минимум простых неорганических соединений: углекислота, самые простые формы азота (аммонийные, нитратные соли или молекулярный азот), минеральные соли (источники фосфора, серы, магния, железа, микроэлементов), вода. Цианобактерии не требуют никаких питательных компонентов в восстановленной форме. Только некоторые морские виды обнаруживают потребность в витамине B12.
Азотфиксирующая активность выявлена более чем у 250 штаммов, принадлежащих к разным группам фототрофных эубактерий. Примерно половину из них составляют цианобактерии. Способность последних к фиксации N2, определяемая по наличию нитрогеназной активности, зависит от условий, и в первую очередь от содержания в среде связанного азота и молекулярного кислорода. Основное место действия обоих факторов — нитрогеназа. В первом случае источники связанного азота репрессируют синтез и ингибируют активность фермента, во втором — O2 быстро инактивирует его.
Отрицательное действие O2 на азотфиксацию связано с восстановительной природой процесса. Возникшая первоначально у анаэробных прокариот, получающих энергию за счет брожения, способность к азотфиксации проявилась и в группах эубактерий с бескислородным фотосинтезом. Благоприятные условия для нее обеспечивались анаэробным типом метаболизма этих групп. И только цианобактерий столкнулись с проблемой функционирования в клетке двух процессов, один из которых имеет восстановительную природу, а другой сопровождается выделением такого сильного окислителя, как O2. Возникла необходимость защиты или изолирования процесса азотфиксации от молекулярного кислорода.
Таблица 28. Способы получения энергии в группе цианобактерий
| Способ получения энергии | Донор электронов | Акцептор электронов | Распространенность и физиологический эффект |
| Фотосинтез кислородного типа | H2O | НАДФ+, ферредоксин | обеспечивает рост всех цианобактерий |
| Фотосинтез бескислородного типа | H2S, Na2S2O3, H2, органические соединения | НАДФ+ ферредоксин | обеспечивает рост некоторых изученных видов; у большинства — снабжает энергией, необходимой для поддержания жизнедеятельности |
| Дыхание | НАД(Ф)-H2 | O2 | обеспечивает рост факультативно фототрофных цианобактерий и поддержание жизнедеятельности облигатно фототрофных видов |
| H2* | O2 | может быть связано с получением энергии | |
| Анаэробное дыхание | НАД(Ф)-H2 | S0 | поддерживает жизнедеятельность некоторых цианобактерий, способных к бескислородному фотосинтезу |
| Брожение | эндогенные или экзогенные сахара | пируват | обнаружено у некоторых факультативно анаэробных цианобактерий; активность недостаточна для поддержания жизнедеятельности** |
* Разные представители цианобактерий оказались способными использовать в темноте молекулярный водород при наличии в качестве акцептора электронов O2. Имеются данные в пользу того, что потребление На связано с функционированием дыхательной цепи и может вести к получению энергии.
** Есть только одно сообщение о способности цианобактерий рода Nostoc расти в темноте в анаэробных условиях, осуществляя сбраживание некоторых сахаров.
Вегетативные клетки многих изученных культур обнаруживают нитрогеназную активность в анаэробных и микроаэробных условиях. Только для единичных культур, например представителей рода Gloeothece, показана способность вегетативных клеток к азотфиксации в аэробных условиях, при этом до 95% фиксированного азота приходится на темновой период, т. е. процессы фотосинтеза и азотфиксации разделены во времени. В целом же проблема фиксации N2 в аэробных условиях значительной частью цианобактерий решена путем формирования дифференцированных клеток определенного типа — гетероцист, в которых чувствительный к O2 аппарат фиксации молекулярного азота отделен от фотосинтетического аппарата с помощью определенных ультраструктурных и биохимических перестроек. Таким образом, способность подавляющего большинства цианобактерий к азотфиксации в аэробных условиях связана с гетероцистами.
При отсутствии в среде связанного азота некоторые вегетативные клетки (обычно 5–10%) нитчатых цианобактерий, принадлежащих к порядкам Nostocales и Stigonematales, превращаются в гетероцисты, образование которых происходит в течение 24 ч параллельно с развитием нитрогеназной активности и может быть разделено на два этапа. Прогетероцисты, формирующиеся на первом этапе, характеризуются неспособностью обеспечить защиту нитрогеназы от инактивирующего действия O2, а также тем, что процесс дифференцировки на этой стадии обратим. На втором этапе процесс дифференцировки становится необратимым. Сформированные гетероцисты не способны к делению и не могут превращаться в вегетативные клетки.
Формирование гетероцист из вегетативных клеток сопровождается глубокими ультраструктурными и функциональными перестройками (рис. 86, А). Зрелые гетероцисты окружены тремя дополнительными слоями, внешними по отношению к клеточной стенке, что затрудняет проницаемость в них воды, ионов, нейтральных веществ гидрофильной природы и растворенных газов. Дополнительные слои, окружающие гетероцисту, в местах ее контакта с вегетативной клеткой прерываются. Перегородка, отделяющая гетероцисту от вегетативной клетки, пронизана множеством мелких каналов (микроплазмодесм), соединяющих цитоплазмы обеих клеток и обеспечивающих обмен клеточными метаболитами. В цитоплазме гетероцист в зонах контакта с вегетативными клетками располагаются светопреломляющие полярные гранулы цианофицина.
Значительную реорганизацию претерпевает в гетероцистах система фотосинтетических мембран: они укорачиваются, теряют расположение, характерное для вегетативных клеток; как правило, отмечается скопление тилакоидов вблизи полюсов гетероцисты. Морфологические изменения тилакоидов сочетаются с важными перестройками фотосинтетического аппарата на функциональном уровне. В гетероцистах не работает II фотосистема. Следовательно, внутриклеточный O2 в них не образуется. Потеря активности II фотосистемы коррелирует со следующими биохимическими особенностями гетероцист: отсутствием основных светособирающих пигментов II фотосистемы — фикобилипротеинов и содержащих их структур — фикобилисом; резко пониженным содержанием ионов марганца — необходимого компонента системы разложения воды; потерей гетероцистами способности фиксировать CO2, связанной с отсутствием рибулозодифосфаткарбоксилазы в растворимой форме или в виде карбоксисом. Деградация II фотосистемы сопровождается сохранением активности I фотосистемы, что находит отражение в поддержании значительного уровня хлорофилла a и увеличении числа реакционных центров этой фотосистемы.
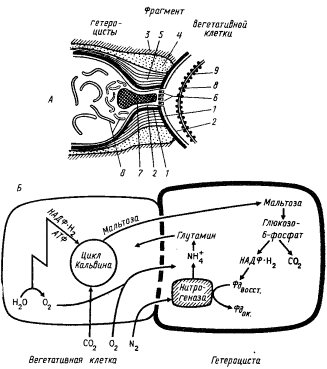 |
| Рис. 86. Схема строения гетероцисты (А) и обмена углеродными и азотными соединениями между гетероцистой и вегетативной клеткой (Б): 1 — клеточная стенка; 2 — ЦПМ; 3 — фибриллярный слой; 4 — гомогенный слой; 5 — пластинчатый слой оболочки гетероцисты; 6 — микроплазмодесмы; 7 — полярная цианофициновая гранула; 8 — тилакоиды; 9 — фикобилисомы |
В процессе формирования гетероцист наблюдается исчезновение различных цитоплазматических включений, характерных для вегетативных клеток: гликогеновых, полифосфатных гранул. В то же время в гетероцистах сохраняется в полном объеме генетическая информация, и в процессе их жизнедеятельности отмечаются активные процессы синтеза РНК и белка. На генетическую "полноценность" гетероцист указывают и неоднократно наблюдавшиеся факты их прорастания и деления.
Для фиксации N2 необходим восстановитель в виде молекул восстановленного ферредоксина (иногда НАДФ-H2) и химическая энергия в форме АТФ. Из-за отсутствия в гетероцистах нециклического транспорта электронов они не могут обеспечивать процесс азотфиксации фотохимически образованным восстановителем и зависят в этом отношении от межклеточного переноса метаболитов (рис. 86, Б). Восстановитель может или непосредственно транспортироваться из соседних вегетативных клеток в готовом виде, или же генерироваться в гетероцистах в темновых ферментативных процессах из исходного транспортируемого субстрата. В последнем случае таким субстратом служит дисахарид мальтоза — продукт восстановительного пентозофосфатного цикла. Ее катаболизирование, осуществляемое по активно функционирующему в гетероцистах окислительному пентозофосфатному пути, приводит к образованию молекул НАДФ-H2, с которых водород может передаваться на ферредоксин в реакции, катализируемой ферредоксин: НАДФ-оксидоредуктазой. Источником АТФ в гетероцистах на свету служит зависимое от I фотосистемы циклическое фотофосфорилирование, в темноте — окислительное фосфорилирование.
Нитрогеназная система катализирует восстановление N2 до аммония. Последний включается в молекулу глутаминовой кислоты в реакции, катализируемой глутаминсинтетазой:
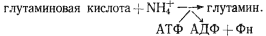
В таком виде фиксированный азот транспортируется из гетероцист в вегетативные клетки, где с помощью глутаматсинтазы осуществляется перенос амидной группы на молекулу -кетоглутарата:
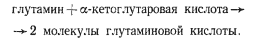
Одна из молекул глутамата возвращается в гетероцисту для очередного акцептирования NH4+, другая поступает в метаболизм вегетативной клетки.
Таким образом, все структурные и функциональные перестройки, происходящие в процессе формирования гетероцист, направлены на поддержание высокой активности нитрогеназы, что достигается, с одной стороны, путем эффективного ее снабжения восстановителем и энергией, с другой — защитой от молекулярного кислорода за счет уменьшения проникновения O2 через утолщенные оболочки гетероцист, реорганизации их фотосинтетического аппарата и высокой активности дыхания.








