 2015-06-24
2015-06-24 577
577В настоящей работе отражены результаты анализа функциональных спектров и таксономической приуроченности белковых доменов, включенных в 15-ую версию системы Pfam (версия опубликована в интернете 20 августа 2004 г.). Данная система, представляющая собой наиболее полный систематизированный каталог такого рода, в настоящее время включает 7503 белковых домена.
Понятие «белковый домен» тесно связано с активно разрабатывающейся в настоящее время естественной классификацией белков. Домен – это более или менее консервативная последовательность аминокислот (или т.н. «мотив» - последовательность, включающая чередующиеся консервативные и вариабельные фрагменты), присутствующая в нескольких (обычно во многих) белковых молекулах у разных организмов. Большинство доменов, включенных в систему Pfam, характеризуются строго определенной функцией и представляют собой, таким образом, функциональные блоки белковых молекул (например, ДНК-связывающие домены или каталитические домены ферментов). Функция некоторых доменов до настоящего времени остается неизвестной, но консервативность и характер распространения этих последовательностей позволяет предполагать, что они также обладают функциональным единством. Предполагается, что подавляющее большинство доменов представляют собой гомологичные последовательности (т.е. имеющие единое происхождение, а не возникавшие параллельно в разных ветвях эволюционного древа). Об этом свидетельствует значительная длина этих последовательностей, а также тот факт, что почти любая функция (каталитическая, сигнальная, структурная и др.) может быть реализована многими разными комбинациями аминокислот, поэтому в случае параллельного возникновения функционально схожих блоков в белковых молекулах у разных организмов факт независимого происхождения, как правило, достаточно очевиден.
|
|
|
Белки объединяются в семейства на основании присутствия в них общих доменов, поэтому понятия «семейство белков» и «домен» в системе Pfam во многом совпадают.
На основе данных из системы Pfam было определено количественное распределение доменов по трем надцарствам живой природы (Archaea, Bacteria, Eukaryota) (рис. 1)
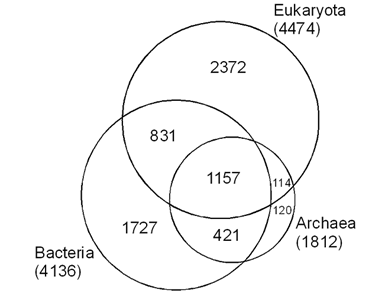
Рис. 1. Количественное соотношение общих и уникальных белковых доменов у архей, бактерий и эвкариот. Площади фигур примерно пропорциональны числу доменов.
Всего в 15-ой версии Pfam присутствует 4474 эвкариотных домена, которые можно подразделить на 4 группы:
1) Специфические домены эвкариот, не встречающиеся в двух других надцарствах (2372);
2) Домены, присутствующие у представителей всех трех надцарств (1157);
3) Домены, общие для эвкариот и бактерий, но отсутствующие у архей (831);
|
|
|
4) Домены, общие для эвкариот и архей, но отсутствующие у бактерий (114).
Наибольшее внимание в последующем обсуждении уделено доменам третьей и четвертой групп, поскольку их таксономическая приуроченность позволяет с определенной долей вероятности говорить об их происхождении. По-видимому, значительная часть доменов третьей группы унаследована эвкариотами от бактерий, четвертой – от архей.
В отдельных случаях общность доменов в разных надцарствах может быть связана с позднейшим горизонтальным переносом, но тогда у надцарства-«реципиента», скорее всего, данный домен будет встречаться лишь у одного или немногих представителей. Такие случаи действительно есть. По сравнению с предыдущей, 14-ой версией Pfam в новой, 15-ой версии целый ряд сугубо бактериальных доменов переместился в третью группу по той причине, что соответствующие последовательности были обнаружены в недавно «расшифрованных» геномах отдельных эвкариот (в особенности комара Anopheles gambiae и простейшего Plasmodium yoelii). Присутствие в геноме малярийного комара генов, кодирующих белки бактериальных жгутиков (притом, что ни у каких других эвкариот эти последовательности не обнаружены), естественно, наводит на мысль о горизонтальном переносе. Подобные домены в дальнейшем обсуждении не учитывались (в третьей группе их около 40, в четвертой группе они отутствуют).
Количественное соотношение общих и уникальных доменов в трех надцарствах, казалось бы, говорит о решительном преобладании в эвкариотической клетке «бактериального» компонента по сравнению с «архейным» (у эвкариот присутствует 831 «бактериальный» домен и 114 «архейных»). Аналогичные результаты получены недавно в ходе сравнительного анализа геномов дрожжей и различных прокариот: оказалось, что 75% от общего числа ядерных генов дрожжей, имеющих прокариотические гомологи, более сходны с бактериальными, чем с архейными последовательностям (Esser et al., 2004). Этот вывод, однако, становится не столь очевидным, если сопоставить упомянутые цифры с суммарным числом общих и уникальных доменов в двух надцарствах прокариот. Так, из общего числа бактериальных доменов, не встреченных у архей (2558), в эвкариотические клетки перешел 831, что составляет 32.5%. Из общего числа архейных доменов, не встречающихся у бактерий (224), в эвкариотических клетках обнаружено 114, то есть 48.7%. Таким образом, если представить себе формирующуюся эвкариотическую клетку как систему, способную к свободному выбору тех или иных белковых блоков из имеющегося набора, то следует признать, что она отдавала предпочтение архейным доменам.
Еще более очевидной становится значительная роль архейного компонента в становлении эвкариот, если сравнить «функциональные спектры» (распределение по функциональным группам) и физиологическое значение эвкариотических доменов «архейного» и «бактериального» происхождения.
Функциональный спектр эвкариотических доменов «архейного» происхождения (таблица; рис. 2).
Первое, что бросается в глаза при просмотре описаний доменов этой группы, это высокая встречаемость таких слов и фраз, как «essential» (ключевой, жизненно важный) и «plays a key role» (играет ключевую роль). В аннотациях доменов из других групп подобные указания встречаются на порядок реже.
В данной группе резко преобладают домены, связанные с самыми базовыми, центральными процессами жизни клетки, а именно с процессами хранения, воспроизведения, структурной организации и считывания генетической информации. Сюда относятся ключевые домены, ответственные за механизм репликации (домены ДНК-примазы и т.п.), транскрипции (включая 7 доменов ДНК-зависимых РНК-полимераз), трансляции (большой набор рибосомных белков, домены, связанные с биогенезом рибосом, факторы инициации и элонгации и т.д.), а также с различными модификациями нуклеиновых кислот (включая процессинг рРНК в ядрышке) и с их организацией в ядре (гистоны и другие белки, связанные с организацией хромосом). Заметим, что недавно проведенный детальный сравнительный анализ всех известных белков, связанных с транскрипцией, показал, что у архей обнаруживается больше сходства с эвкариотами, чем у бактерий (Coulson et al., 2001, fig.1b).
|
|
|
Представляют интерес 6 доменов, связанных с синтезом (посттранскрипционными модификациями) тРНК. Химические изменения, вносимые специальными ферментами в нуклеотиды тРНК, являются одним из важнейших средств адаптации к высоким температурам (они позволяют тРНК сохранять правильную третичную структуру при нагревании). Показано, что число измененных нуклеотидов в тРНК термофильных архей растет при повышении температуры (Noon et al., 2003). Сохранение этих архейных доменов у эвкариот, возможно, указывает на то, что температурные условия в местообитаниях первых эвкариот были нестабильными (существовала опасность перегрева), что характерно для мелководных местообитаний.
Сигнально-регуляторных доменов сравнительно немного, но среди них – такие важные, как транскрипционный фактор TFIID (TATA-связывающий белок, PF00352), домены транскрипционных факторов TFIIB, TFIIE, TFIIS (PF00382, PF02002, PF01096), - регуляторы транскрипции общего назначения, играющие центральную роль в активации генов, транскрибируемых РНК-полимеразой II. Интересен также домен CBFD_NFYB_HMF (PF00808): у архей это гистон, а у эвкариот – гистоноподобный транскрипционный фактор.
Особо следует отметить эвкариотные домены «архейного происхождения», связанные с мембранными пузырьками. К ним относится домен Adaptin N (PF01602), связанный у эвкариот с эндоцитозом; Aromatic-di-Alanine (AdAR) repeat (PF02071), у эвкариот участвующий в процессе слияния мембранных пузырьков с цитоплазматической мембраной и обнаруженный у двух видов архей из рода Pyrococcus; Syntaxin (PF00804), у эвкариот регулирующий, в частности, присоединение внутриклеточных мембранных пузырьков к пресинаптической мембране нейронов и обнаруженный у аэробных архей рода Aeropyrum, и др. Среди «доменов бактериального происхождения» белки с такими функциями отсутствуют. Домены, управляющие слиянием мембран и образованием пузырьков, могли сыграть важную роль в симбиогенетическом становлении эвкариотической клетки, поскольку создают базу для развития фагоцитоза (наиболее вероятный путь приобретения внутриклеточных симбионтов – пластид и митохондрий), а также для слияния клеток (копуляции) и образования различных внутриклеточных мембранных структур, свойственных эвкариотам, таких как эндоплазматический ретикулум (ЭР). ЭР эвкариот, согласно одной из гипотез, имеет архебактериальное происхождение (Dolan et al., 2002). Предположение основывается, в частности, на сходстве синтеза N-связанных гликанов в ЭР с определенными этапами формирования клеточной стенки у архей (Helenius, Aebi, 2001). Напомним, что ЭР эвкариот тесно связан с ядерной оболочкой, что позволяет предполагать единый генезис этих структур.
|
|
|
Следует обратить внимание также на практически полное отсутствие в этой группе метаболитических доменов (что представляет резкий контраст с группой эвкариотных «доменов бактериального происхождения», где метаболитические белки, напротив, резко преобладают).
С точки зрения проблемы возникновения эвкариот представляют интерес такие домены архейного происхождения, как ZPR1 zinc-finger domain (PF03367) (у эвкариот этот домен входит в состав множества ключевых регуляторных белков, особенно тех, которые отвечают за взаимодействие между ядерными и цитоплазматическими процессами), и zf-RanBP (PF00641), являющийся у эвкариот одим из важнейших компонентов ядерных пор (отвечает за транспорт веществ через ядерную мембрану).
Все 28 доменов рибосомальных белков архейного происхождения присутствуют в составе цитоплазматических рибосом эвкариот, причем все они обнаруживаются как у растений, так и у животных. Этой картине хорошо соответствует тот факт, что домен NOG1, обладающий специфической ГТФ-азной активностью и используемый вспомогательными белками ядрышкого организатора (кластеры генов рРНК) также имеет архейное происхождение.
Таблица. Сравнение функциональных спектров эвкариотических доменов, имеющихся или отсутствующих у архей (А), цианобактерий (Ц), альфапротеобактерий (П) и бактерий в целом, включая Ц и П (Б).
| Функциональная группа | Есть у А, нет у Б | Есть у Б, нет у А | Есть у Ц или П, нет у А | Есть у Б, нет у А, Ц и П |
| Синтез белка | ||||
| В том числе: рибосомные и связанные с биогенезом рибосом | ||||
| Трансляция | ||||
| Синтез, модификация тРНК | ||||
| Посттрансляционные модификации белков | ||||
| Репликация, транскрипция, модификация и организация НК | ||||
| В том числе: базовая репликация и транскрипция | ||||
| Гистоны и другие белки, организующие ДНК в хромосомах | ||||
| Модификация НК (нуклеазы, топоизомеразы, геликазы и т.д.) | ||||
| Репарация, рекомбинация | ||||
| НК-связывающие домены неясной функции или общего назначения | ||||
| Белки, связанные с образованием и функционированием мембранных пузырьков | ||||
| Транспортные и сортировочные белки | ||||
| Сигнальные и регуляторные белки | ||||
| В том числе: транскрипционные факторы (регуляция экспрессии генов) | ||||
| Рецепторы | ||||
| Домены межклеточного взаимодействия | ||||
| Домены межбелкового взаимодействия | ||||
| Домены связи белка с мембраной | ||||
| Защитные и связанные с иммунной системой | ||||
| Связанные с вирулентностью патогенных бактерий и простейших | ||||
| Регуляция онтогенеза | ||||
| Домены, связанные с гормонами | ||||
| Токсины | ||||
| Апоптоз | ||||
| Регуляция репликации | ||||
| Лектины (белки, образующие комплексы с углеводами) | ||||
| Прочие сигнальные и регуляторные белки | ||||
| Белки, связанные с цитоскелетом, микротрубочками | ||||
| Белки, связанные с клеточным делением | ||||
| Метаболизм | ||||
| В том числе: кислородное окисление (оксигеназы, пероксидазы и т.п.) | ||||
| Метаболизм стероидов, терпенов | ||||
| Метаболизм нуклеотидов и азотистых оснований | ||||
| Метаболизм углеводов | ||||
| Метаболизм липидов | ||||
| Метаболизм аминокислот | ||||
| Метаболизм белков (пептидазы, протеазы и пр.) | ||||
| Фотосинтез, дыхание, цепь переноса электронов | ||||
| Прочая базовая энергетика (АТФ-синтазы, НАД-Н дегидрогеназы и пр.) | ||||
| Другие метаболитические домены |
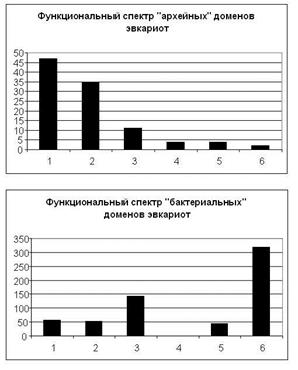
Рис. 2. Функциональные спектры «архейных» и «бактериальных» доменов эвкариот. 1 – Синтез белка, 2 - Репликация, транскрипция, модификация и организация НК, 3 - Сигнальные и регуляторные белки, 4 - Белки, связанные с образованием и функционированием мембранных пузырьков, 5 - Транспортные и сортировочные белки, 6 – Метаболизм








