 2015-06-26
2015-06-26 1734
1734Тип I строения клеточных стенок характерен для большинства двудольных и «некоммелиноидных» однодольных. Клеточные стенки этого типа содержат относительно равные количества целлюлозы и ксилоглюканов. XyGs прочно связываются с поверхностными глюкановыми цепочками в целлюлозных микрофибриллах и фиксируют микрофибриллы в нужном пространственном положении, связываясь не только с целлюлозой, но и между собой. При этом они заполняют пространство между смежными целлюлозными микрофибриллами.
В клеточной стенке первого типа Каракас, состоящий из целлюлозы и XyGs, помещен в матрикс пектинов, который регулирует, помимо других физиологических свойств, размер пор клеточной стенки. У гомогалактоуронанов (HGAs), секретируемых из цитоплазмы, большинство карбоксильных групп метилировано, т.е. HGAs попадают в клеточную стенку в виде высокометоксилированных полимеров. В клеточной стенке работает фермент пектинметилэстераза (РМЕ), которая отщепляет некоторые из метальных групп. Освободившиеся карбоксильные группы могут соединяться с ионами Са2+ . Кальций как
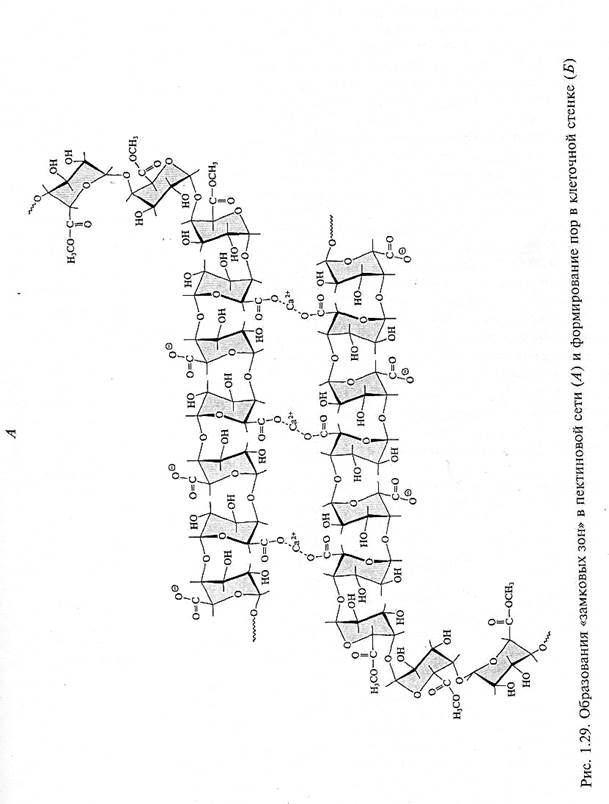
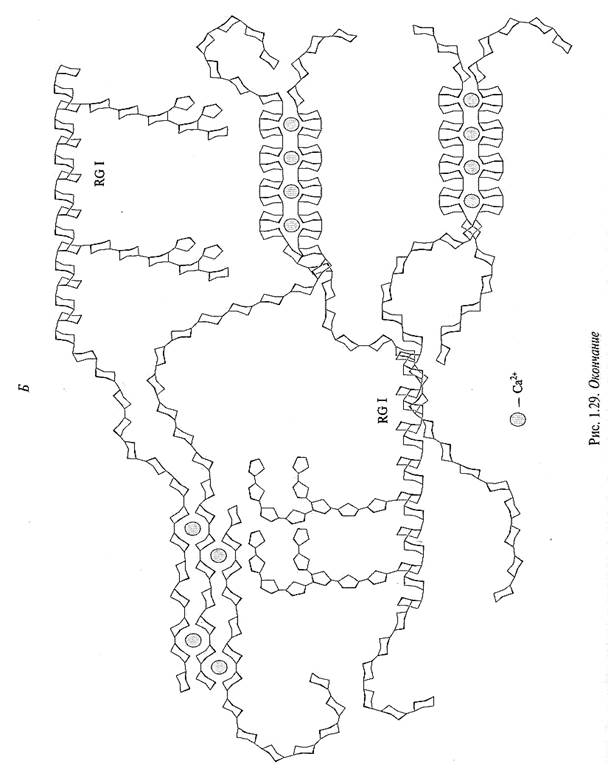
двухвалентный катион может связываться с двумя карбоксильными группами двух цепей пектинов. При этом образуются «замковые зоны», соединяющие две антипараллельные цепи HGAs. Наиболее сильные связи между цепями HGAs образуются, если в «замковой зоне» расположены не менее семи неэтерифицированных остатков галактуроновых кислот. Полагают, что от расстояния между «замковыми зонами» зависит, какого размера поры образуют в клеточной стенке несвязанные участки HGAs: чем больше расстояние между «замковыми зонами», тем больше размер пор (рис. 1.29).
Наряду с гомогалактоуронанами в формировании пор клеточных стенок принимают участие и рамногалактоуронаны I (RG I). Во-первых, их рамнозные остатки в основной цепи и боковые цепочки мешают образованию прочных «замковых зон». Во-вторых, нейтральные полимеры (арабинаны или галактаны), прикрепленные одним концом к RG I, имеют много степеней свободы и достаточно подвижны. Они могут располагаться внутри пор, уменьшая их размеры (рис. 1.29). На некоторых стадиях развития клетки значительно повышается активность специфичных гидролаз, при этом нейтральные полимеры отщепляются и размер пор увеличивается. Другие компоненты пектиновой сети также могут участвовать в регулировании «порозистости» клеточной стенки.
Тип II строения клеточных стенок характерен для «коммелиноидных» однодольных. Подобно типу I он содержат микрофибриллы целлюлозы, однако они соединяются между собой не ксилоглюканами, а глюкуроноарабиноксиланами (GAXs). Боковые цепочки у основной ксилановой цепи GAXs по-разному влияют на образование связей. Ответвления, состоящие из L-арабинозы и L-глюкуроновой кислоты, предотвращают формирование водородных связей и блокируют перекрестное сшивание между полисахаридами. Напротив, D-ксилозные единицы стабилизируют линейную структуру полимера и способствуют его связыванию с глюканами.
Клеточные стенки второго типа достаточно бедны пектинами. Частично функции пектинов могут выполнять остатки L-галактуроновах кислот, входящих в состав GAXs. Порозистость и прочность клеточной стенки зависит от интервала между боковыми Ага и GlcA единицами GAXs, которые таким образом функционально заменяют пектиновые вещества клеточных стенок первого типа.
И наконец, клеточные стенки типа II имеют очень немного структурных белков по сравнению с двудольными и другими однодольными растениями. В то же время в них часто формируются обширные сети фенилпропаноидных соединений, особенно после прекращения роста клетки.

