 2015-07-04
2015-07-04 2744
2744Основными светочувствительными элементами (рецепторами) являются два вида клеток: одни в виде стебелька - палочки 110-123 млн. (высота 30 мкм, толщина 2мкм), другие более короткие и более толстые - колбочки 6-7 млн. (высота 10мкм, толщина 6-7 мкм). Они распределены в сетчатке неравномерно. Центральная ямка сетчатки(fovea centralis) содержит только колбочки(до 140 тыс. на 1 мм 52 0). По направлению к периферии сетчатки их число уменьшается, а число палочек возрастает.
Каждый фоторецептор - палочка или колбочка состоит из чувствительного к действию света наружного сегмента содержащего зрительный пигмент и внутреннего сегмента, который содержит ядро и митохондрии обеспечивающие энергетические процессы в фоторецепторной клетке
{ Наружный сегмент светочувствительный участок, где световая энергия преобразуется в рецепторный потенциал Электронно-микроскопические исследования выявили, что наружный сегмент заполнен мембранными дисками, образованными плазматической мембраной. В палочках, в каждом наружном сегменте, содержится 600-1000 дисков, которые представляют собой уплощенные мембранные мешочки, уложенные как столбик монет. В колбочках мембранных дисков меньше. Это частично объясняет более высокую чувствительность палочки к свету (палочку может возбудить всего один квант света, а для активации колбочки требуется больше сотни квантов).
Каждый диск представляет собой двойную мембрану, состоящую из двойного слоя молекул фосфолипидов, между которыми находятся молекулы белка. С молекулами белка связан ретиналь, входящий в состав зрительного пигмента родопсина.
Наружный и внутренний сегменты фоторецепторной клетки разделены мембранами, через которые проходит пучок из 16-18 тонких фибрил. Внутренний сегмент переходит в отросток, с помощью которого фоторецепторная клетка передает возбуждение через синапс на контактирующую с ней биполярную нервную клетку}
Наружные сегменты рецепторов обращены к пигментному эпителию, так что свет в начале проходит через 2 слоя нервных клеток и внутренние сегменты рецепторов, а потом достигает пигментного слоя.
Колбочки функционируют в условиях больших освещенностей - обеспечивают дневное и цветовое зрение, а палочки - отвечают за сумеречное зрение.
Видимый нами спектр электромагнитных излучений заключен между коротковолновым (длина волны от 400нм) излучением, которое мы называем фиолетовым цветом и длинноволновым излучением(длина волны до 700 нм) называемым красным цветом. В палочках находится особый пигмент- родопсин, (состоит из альдегида витамина А или ретиналя и
белка) или зрительный пурпур, максимум спектра, поглощения которого находится в области 500 нанометров. Он ресинтезируется в темноте и выцветает на свету. При недостатке витамина А нарушается сумеречное зрение -"куриная слепота".
В наружных сегментах трех типов колбочек(сине-, зелено и красно-чувствительных) содержится три типа зрительных пигментов, максимум спектров поглощения которых находится в синей (420 нм), зеленой(531 нм) и красной(558 нм) частях спектра. Красный колбочковый пигмент получил название - " йодопсин". Структура йодонсина близка к родонсину.
{ Рассмотрим последовательность изменений:}
Молекулярная физиология фоторецепции: Внутриклеточные регистрации от колбочек и палочек животных показали, что в темноте вдоль фоторецептора течет темновой ток, выходящий из внутреннего сегмента и входящий в наружный сегмент. Освещение приводит к блокаде этого тока. Рецепторный потенциал модулирует выделение медиатора (глутамата) в синапсе фоторецептора. Было показано, что в темноте фоторецептор непрерывно выделяет медиатор, который действует деполяризующим образом на мембраны постсинаптических отростков горизонтальных и биполярных клеток.(см. Рис.1)
Палочки и колбочки обладают уникальной среди всех рецепторов электрической активностью, их рецепторные потенциалы при действии света - гиперполяризующие, потенциалы действия под их влиянием не возникают.
{ При поглощении света молекулой зрительного пигмента.(см. Рис.2) - родопсина в ней происходит мгновенная изомеризация ее хромофорной группы: 11- цис -ретиналь превращается в транс -ретиналь. Вслед за за фотоизомеризацией ретиналя происходят пространственные изменения в белковой части молекулы: она обесцвечивается и переходит в состояние метородопсина II. В результате этого молекула зрительного пигмента приобретает способность к взаимодействию с другим примембранным белком г уанозин трифосфат(ГТФ) - связывающим белком – трансдуцином (Т).
Рис.1 Рис 2. 
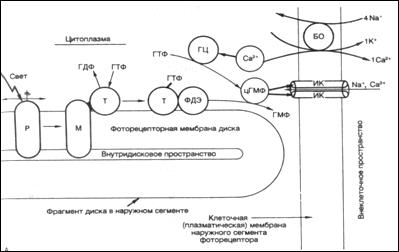
В комплексе с метародопсином трансдуцин переходит в активное состояние и обменивает связанный с ним в темноте ганозитдифосфат(ГДФ) на (ГТФ). Трансфдуцин + ГТФ, активируют молекулу другого примеммбранного белка - фермента фосфодиэстеразы(ФДЭ). Активированная ФДЭ разрушает несколько тысяч молекул цГМФ.
В результате падает концентрация цГМФ в цитоплазме наружного сегмента рецептора. Это приводит к закрытию ионных каналов в плазматической мембране наружного сегмента, которые были открыты в темноте и через которые внутрь клетки входили Na + и Ca. Ионные каналы закрываются вследствие того, что падает концентрация цГМФ, которая держала каналы открытыми. В настоящее время выяснено, что поры в рецепторе открываются благодаря цГМФ циклическому гуанозинмонофосфату.
Механизм восстановления исходного темного состояния фоторецептора связан с повышением концентрации цГМФ. (в темновую фазу с участием алкагольдегидрогеназы+ НАДФ)
Т.о поглощение света, молекулами фотопигмента приводит к снижению проницаемости для Nа, что сопровождается гиперполяризацией, т.е. возникновению рецепторного потенциала.}
Гиперполяризационный рецепторный потенциал, возникший на мембране наружного сегмента, распространяется затем вдоль клетки до ее пресинаптического окончания и приводит к уменьшению скорости выделения медиатора - глутамата.








