 2015-07-03
2015-07-03 1902
1902Транспирация. Восходящий водный ток заканчивается испарением в процессе транспирации подавляющего количества воды, поступающей из почвы через корень. Транспирация – это процесс испарения воды надземными органами растения. Расходование воды растением регулируется целым рядом анатомических и физиологических организмов. Поэтому транспирация является физиологическим процессом в отличии от чисто физического испарения.
Биологическое значение транспирации состоит в терморегуляции растения, обеспечении деятельности верхнего концевого двигателя водного тока, при помощи которого осуществляется и поступление различных веществ, а также в регулировании насыщенности клеток водой, благодаря чему создаются оптимальные условия для процессов жизнедеятельности.
Одной их важных характеристик процесса является интенсивность транспирации – количество воды, испаряемое растением с единицы листовой поверхности в единицу времени. Иногда расчет ведут не на единицу площади. А на единицу массы листьев. Испарение во много раз превышает необходимый для растений расход воды. Так наиболее пышное развитие растений наблюдается во влажных тропиках, а также в оранжереях, где влажность воздуха высокая, а транспирация низкая. Кроме того, известна положительная роль полива дождеванием, способствующего транспирации. У большинства сельскохозяйственных растений интенсивность транспирации колеблется от 15 до 250 г воды/м2∙ч, а ночью может снижаться до 7–20 г воды/м2∙ч и даже больше. На гектар пашни культурные растения в дневные часы в среднем расходуют 100 т воды. Высокая интенсивность транспирации, которую К.А. Тимирязев назвал «необходимым злом», обусловлена тем, что атмосфера характеризуется очень низкими значениями водного потенциала. Водный потенциал связан логарифмической зависимостью с относительной влажностью воздуха
Ψ = 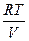 ∙ ln
∙ ln 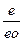 ,
,
где е – давление водяного пара в воздухе;
е0 – давление водяного пара, насыщающего воздух при данной температуре.
Поэтому небольшой перепад относительной влажности воздуха приводит к значительной депрессии его водного потенциала. Так, если при 100 % относительной влажности воздуха водный потенциал равен 0, то при влажности 99,6 % - (-0,5) МПа, а при 99 и 97 % соответственно –1,36 и – 4,0 МПа. Часто летом относительная влажность воздуха равна 60%, водный потенциал при этом снижается до -68 МПа, а во время суховея (влажность 30 %) депрессия водного потенциала достигает -200 МПа.
С самого начала существования наземных растений возникла дилемма: ассимиляция СО2 из атмосферы требует интенсивного газообмена, а предотвращение значительной потери воды возможно только при наличии хорошей изоляции от окружающего воздуха, имеющего крайне низкие значения водного потенциала. Главная проблема газообмена в растении, по мнению О. Штоккера, «лавирование между жаждой и голодом».
Поддержание водного гомеостаза листьев и молодых стеблей достигается наличием покровной ткани – эпидермиса, который состоит из одного слоя клеток, наружные стенки которых утолщены. Кутикула является внешним защитным покрытием клеток эпидермиса. Она защищает растения не только от потери воды, но также является барьером для бактерий и грибов. В состав кутикулы входит кутин и воска. Кутин – это полимер, состоящий из гидроксимонокарбоновых кислот, содержащих 16-18 атомов углерода и гидроксильные группы. Эти кислоты соединены в цепочки с помощью эфирных связей. Воска не являются полимерами. Это смесь алканов, жирных кислот и спиртов и их эфиров. Они обладают высокогидрофобными свойствами. Воска синтезируются в эпидермальных клетках, часто кристаллизуются.
Кутикула – трехслойная структура: 1) слой воска; 2) истинный кутин, погруженный в воск; 3) кутикулярный слой, включающий кутин, воска и углеводороды, которые смешиваются с элементами клеточной стенки.
Кутикулярное диффузионное сопротивление в большинстве случаев очень велико. Оно зависит от толщины кутикулы, расположения, плотности и числа прослоек кутина и воска. На формирование кутикулы сказываются видовые особенности и возраст растений, а также условия их произрастания. При засухе у взрослых листьев более толстая кутикула и более плотный восковой слой. Кутикулярная защита от транспирации весьма эффективна. У взрослых листьев кутикулярная транспирация составляет 10-20% от общего испарения воды.
У кутикулы есть уникальное свойство, обусловленное её составом, - изменять гидравлическую проводимость в зависимости от оводненности. При подсыхании наружных стенок эпидермиса гидрофобные слои кутикулы уплотняются, поэтому кутикулярное сопротивление может удваиваться. При низких температурах оно также возрастает. И, наоборот, при увеличении оводненности эпидермиса кутикула набухает за счет гидратации карбоксильных и оксигрупп, разрыхляется. При этом кутикулярное сопротивление диффузии значительно снижается, и транспирация возрастает. Таким образом, потеря воды через кутикулу регулируется оводненностью листа. По ночам кутикулярная транспирация идет интенсивнее, чем днем.
Устьичная транспирация происходит через устьица. Устьица образованы двумя специализированными эпидермальными клетками, которые называют замыкающими. Между ними находится устьичная щель. Замыкающие клетки способны изменять свою форму, вызывая открытие или закрытие устьичной щели. Вокруг замыкающих клеток имеются чаще всего побочные клетки, которые способны принимать участие в движении устьиц. Количество устьиц варьирует в зависимости от возраста листа от 50 до 500 на 1 мм2.
Отличительной особенностью замыкающих клеток является строение их клеточной стенки. Толщина её неравномерна. У двудольных – она толще в вентральной части клеток, которая граничит с устьичной щелью. Это имеет значение в изменении формы замыкающих клеток в ответ на изменение тургорного давления. Важная роль в регуляции формы замыкающих клеток принадлежит также микрофибриллам целлюлозы, радиально ориентированным в клеточной стенке. Они выполняют роль своеобразных направляющих при движении устьиц.
Несмотря на то, что площадь устьичных отверстий по отношению к поверхности листа составляет всего около 1 %, испарение воды через них более интенсивно. Это объясняется тем, что скорость диффузии газов через малые отверстия пропорциональна не площади, а диаметру отверстия, то есть периметру.
Транспирация – это верхний концевой двигатель водного тока. Этот двигатель развивает силу от -1,0 до -1,5МПа и более, а нижний – от -0,15 до -0,3 МПа.
Природа верхнего концевого двигателя восходящего водного тока. 1. Традиционные представления о присасывающем действии транспирации как единственном верхнем концевом двигателе. Традиционно считается, что неизбежному испарению воды способствуют: 1) резкий перепад водного потенциала между листом и окружающим воздухом; 2) отсутствие у растений надежных покровов, защищающих их от больших потерь воды. Также установилось мнение, что транспирация осуществляется целиком за счет солнечной энергии и не связана с потреблением метаболической энергии (энергии дыхания). Транспирация обладает четко выраженным присасывающим действием, что создает пронизывающую все растение гидростатическую тягу. Транспирационная тяга, благодаря непрерывности водной фазы в растении и силам сцепления молекул воды, буквально вытягивает воду из растения, выполняющего роль соединительного канала между почвой и атмосферой (рис. 15). Компоненты этой системы имеют следующий водный потенциал: почвенный раствор – (-0,05) МПа, корневой волосок – (-0,09) МПа, корень (первичная кора) – (-0,2) МПа, стебель – (-0, 5) МПа, лист – (-1,5) МПа, воздух (при относительной влажности 50 %) – (-100) МПа. Вода в листе из трахеидных окончаний проводящих пучков направляется по апопласту (клеточным стенкам) к местам испарения – предустьичному пространству (дворику, замыкающим клеткам устьиц или эпидермису – кутикулярная транспирация). Передвижение по апопласту – путь наименьшего сопротивления.
2. Возможность восходящего водного тока при подавленной транспирации. Открытие «биогенного» насоса. Хотя трудно отрицать роль присасывающего действия транспирации в подъёме воды по растению, всё же получено не мало фактов, не укладывающихся в рамки описанных представлений. Так, у полностью погруженных в воду растений циркуляция воды осуществляется без всякой транспирационной тяги. При 100 % относительной влажности, когда транспирация подавлена, отсутствует градиент водного потенциала между листом и окружающим воздухом. В этих условиях восходящий ток должен остановиться, однако он присутствует в наземных растениях. В опытах В.Г. Реуцкого и П.А. Родионова с ячменем показано, что при насыщающей влажности воздуха прослеживался как сам поток жидкости по стеблю, так и его температурная зависимость. Q10 скорости движения жидкости составлял 1,27-1,30 в интервале температуры 20-30ºС при 100 % влажности воздуха. В отдельных случаях движение не прекращалось даже при полном погружении листа в воду. Поток жидкости был светозависим. В темноте скорость его передвижения оказалась более чем в два раза ниже, чем на свету. Авторы сделали заключение о существовании системы транспорта воды, не связанной с транспирационной активностью. Светочувствительность транспорта послужила основанием для предположения об участии фотосинтетических реакций в работе верхнего концевого двигателя. С этой целью лист инфильтрировали диуроном – ингибитором фотосинтеза. А затем измеряли скорость потока воды по стеблю при 20 ºС и насыщенной влажности воздуха (транспирация отсутствовала). Под действием диурона скорость потока уменьшилась на 48 % за сутки, а через 6 суток – на 63 %. Авторы сделали вывод, что в механизме передвижения воды принимают участие фотохимические процессы фотосинтеза. Однако они не отрицали и вклад градиента водного потенциала между листом и воздухом в движущую силу транспорта воды. Ими было высказано предположение о двухкомпонентности движущей силы транспорта воды: «пассивного» и «биогенного» насосов. «Пассивный» насос – это присасывающее действие транспирации. Его силой является градиент водного потенциала между листом и атмосферой, а регулярной основой – изменения апертуры устьиц. «Биогенный» насос в чистом виде проявляется при 100 % влажности воздуха, когда транспирация подавлена. Его работа в значительной степени детерминируется фотозависимой системой листа, так как свет может индуцировать увеличение осмотического потенциала клеток листа, а значит уменьшать их водный потенциал, способствуя притоку к ним воды.
А.А. Зялалов и В.Н. Лобанов, проводившие свои опыты также при 100 % влажности воздуха, но не на свету, а в темноте, в качестве возможного эндогенного источника энергии для транспорта воды рассматривали дыхание, с чем нельзя не согласиться.
3. Существует ли постоянно в сосудах ксилемы непрерывный градиент отрицательного гидростатического давления? Немецкий биофизик У. Циммерман при работе со специальным зондом ксилемного давления неожиданно обнаружил в сосудах ксилемы не отрицательное, а, напротив, положительное гидростатическое давление. Это вызвало живейший отклик. Вскоре появились работы, вновь подтверждающие существование в сосудах отрицательного гидростатического давления.
Приведенные факты свидетельствуют о более сложной природе верхнего концевого двигателя. Об этом свидетельствует и автоколебательный характер как транспирации, так и поступления в лист воды и, самое главное, асинхронность осцилляций этих двух процессов. Из которых складывается водообмен листа.
Водообмен листа оказался идентичным водообмену корня. В корне обнаружены находящиеся в противофазе короткопериодные автоколебания поглощения воды и выделения экссудата, в листе же – находящиеся в противофазе короткопериодные автоколебания транспирации и поступления в лист воды.
Наиболее обстоятельно кинетика водообмена листа прослежена в работах Д.В. Шерифа. Он зарегистрировал находящиеся в противофазе короткопериодные (от 1 до 10 мин.) автоколебания транспирации и поступления воды в лист у разных видов растений при работе как с отделенными, так и с неотделенными листьями. Причем эти автоколебания не были связаны с изменением апертуры устьиц.
Противофазность короткопериодичных автоколебаний отмечали и другие авторы. Так, в опытах В.Ю. Ушакова и И.Р. Колтуновой период осцилляции составлял около 6 мин. (рис. 16). Под действием анастетика – хлороформа - автоколебания полностью исчезали, хотя процессы водообмена и продолжались, но с пониженной скоростью.
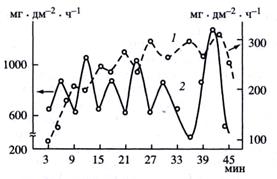 |
Рис. 16. Автоколебания водопоглощения (1) и транспирации (2) у листьев Vicia faba.
В опытах В.Н. Жолкевича и М.Д. Кушниренко с интактными растениями испытано влияние на кинетику процессов водообмена листа некоторых метаболических регуляторов (2.4-динитрофенола, колхицина, гибберелловой кислоты). Ингибиторы ослабляли автоколебания, уменьшая их амплитуду, увеличивая период и уменьшая противофазность. Гиббереловая кислота вызывала противоположный эффект. Автоколебания транспирации и поступления в лист воды и особенно их противофазность оказались зависимы от энергоснабжения, а также чувствительны к дезинтегратору микротрубочек, то есть зависят от функционирования контрактильных систем.
По-видимому, речь снова идет о двух составляющих: метаболической (связанной с автоколебаниями и с передвижением воды по симпласту) и «физической» (не связанной с автоколебаниями, когда вода передвигается по апопласту).
Об ограниченности традиционных представлений о транспирационной тяге как единственном верхнем концевом двигателе говорит и тот факт, что наряду с парообразной водой из листа выделяется и капельно-жидкая вода (не гута). С.А. Кубичек обнаружил на поверхности устьичных замыкающих клеток капельки воды при просматривании нижнего эпидермиса листьев щавеля, лука, традесканции под люминесцентным микроскопом в сине-фиолетовых лучах. При усилении освещения темп выделения капелек возрастал. Д.И. Колпаков обнаружил такой же эффект, используя световой микроскоп. Факты секреции капельно-жидкой воды, причем имеющей пульсирующий характер, является серьёзным аргументом в пользу сложной природы транспорта воды в листе.

