 2015-07-03
2015-07-03 2691
2691Осмотическая концепция экссудации и её ограниченность. Согласно этой концепции, роль находящегося внутри осмометра концентрированного раствора отводится заполняющему проводящие сосуды ксилемному соку, а роль полупроницаемой перегородки, препятствующей выходу наружу ксилемного сока, но пропускающей внутрь воду – эндодерме в совокупности со всеми паренхимными клетками.
Таким образом, выходит, что воду насасывает лишь ксилемный сок, а живые клетки (именно для которых вода и поступает в растение) активного участия в транспорте не принимают. Экссудация при этом подчиняется следующему уравнению:
Jw = (δi∙ОРi – δl∙ОРl)∙Lp,
где Jw – объем водного потока;
ОРi, ОРl – осмотическое давление ксилемного сока и наружного раствора соответственно;
δi, δl – коэффициенты отражения для растворенных веществ ксилемного сока и наружного раствора;
Lp – гидравлическая проводимость корня,
или Jw = δ∙∆ОР∙Lp.
В случае справедливости осмотической концепции, экссудация у отделенной корневой системы должна останавливаться уже в изотоническом по отношению к ксилемному соку растворе (то есть когда ОРi= ОРl), а в гипертонических растворах она вообще невозможна. Однако выяснилось, что экссудация продолжается в заведомо гипертонических по отношению к экссудату растворах, прекращаясь лишь тогда, когда ОРl в среднем вдвое превышает ОРi.
Впервые обнаруживший подобное явление в 1942 году голландский исследователь Д. Овербик сделал заключение о суммировании корневого давления из двух составляющих – осмотической и неосмотической, которую он назвал «активным» давлением. Последнее в его опытах равнялось 48-71 % от осмотического давления раствора, останавливавшего экссудацию, и обратимо подавлялось КСN, то есть оно оказалось энергозависимым.
Существование неосмотической составляющей корневого давления позднее было подтверждено в целом ряде работ. Особенно обстоятельные исследования выполнены Л.В. Можаевой и Н.В. Пильщиковой, которые впервые высказали предположение об участии контрактильных белков в нагнетающей деятельности корня и привели конкретные факты, подтверждающие справедливость такого предположения.
Неосмотическую составляющую стали называть метаболической вследствие её чувствительности к действию метаболических регуляторов. Осмотическое давление наружного раствора, при котором экссудация прекращается, названо компенсационным давлением. Его принимают равным движущей силе экссудации, то есть корневому давлению. Компенсационный способ определения величины корневого давления предложен Д.А. Сабининым (1923), а несколько позднее Н.А. Максимовым.
При учете сложной природы корневого давления, суммирующегося из осмотической и метаболической составляющей, уравнение экссудации будет выглядеть:
Jw = δ∙∆ОР∙Lp + МС,
где δ∙∆ОР∙Lp – осмотическая составляющая,
МС – метаболическая составляющая.
Величина последней достигает 80 % и более от компенсационного давления.
Существуют и другие доказательства недостаточности одних только осмотических критериев для объяснения природы движущих сил транспорта воды. А.П. Петровым установлен факт аддитивности корневого давления. Он установил, что чем длиннее корень, тем больше корневое давление он развивал (табл. 3).
Таблица 3. Зависимость корневого давления от длины зародышего корня Zea mays L. (по А.П. Петрову, 1974)
| Длина корня, см | Давление, см. водного столба | Длина корня, см | Давление, см. водного столба | Длина корня, см | Давление, см. водного столба |
| 15,5 | 140,0 | 9,5 | 117,0 | 4,5 | 40,0 |
| 12,2 | 130,0 | 6,5 | 90,0 |
Иными словами, чем больше количество клеток участвовало в нагнетании воды, тем большим оказывалось и корневое давление. Если бы корневое давление определялось прежде всего градиентом осмотического давления между ксилемным соком и наружным раствором, то оно не зависело бы от длины корня.
Об аддитивности корневого давления могут свидетельствовать и опыты с частичной обрезкой корневой системы яблони (Пильщикова, 1982). При удалении 1/3 корней интенсивность экссудации и метаболическая составляющая корневого давления уменьшались на 30 %. По мере последующего регенерирования интенсивность экссудации и метаболическая составляющая возрастали в точном соответствии с размерами корневой системы. Таким образом, аддитивность является индикатором участия живых клеток в формировании корневого давления и, прежде всего, его метаболической составляющей.
Природа метаболической составляющей корневого давления. Чтобы её определить, было испытано воздействие различных метаболических регуляторов как ингибиторов, так и стимуляторов. Метаболическая составляющая оказалось очень лабильной. При различных воздействиях она варьировала от нескольких процентов до 80 % и более от величины суммарного корневого давления (Жолкевич и др., 1979).
Подтверждена установленная Овербиком энергозависимость метаболической составляющей. Это было показано в опытах с разобщителем окисления и фосфорилирования – 2,4-динитрофенолом. Он уменьшал её.
Метаболическая составляющая чувствительна к релаксанту мышц d-тубокурарину, то есть блокатору контрактильных систем, лишающему животных способности к движению.
Метаболическая составляющая оказалась чувствительной и к нейромедиатору животных ацетилхолину, передающему возбуждение с нервных окончаний на мышцу. Он заметно увеличивал как метаболическую составляющую, так и суммарное корневое давление.
Кальций («запускающий» работу миозина) увеличивает метаболическую составляющую вдвое, а корневое давление на 50 % по сравнению с контролем. Агенты, связывающие или вытесняющие кальций (ЭДТА, пипольфен), оказывали прямо противоположное действие.
Исключительно важным фактом является то, что при всех перечисленных воздействиях суммарное корневое давление и метаболическая составляющая изменяются в одном и том же направлении, тогда как осмотическое давление экссудата (принимаемое за максимальную величину осмотической составляющей) – в противоположном. Изменение скорости экссудации положительно коррелирует с изменением метаболической составляющей и отрицательно – с изменением осмотического давления экссудата. Все это указывает на главенствующую, решающую роль метаболической составляющей в создании корневого давления. Поэтому транспорт воды в корне должен происходить в основном от клетки к клетке, то есть по симпласту, а не только и не столько по апопласту (как полагали и до сих пор все ещё полагают некоторые физиологи). Вероятно, как уже отмечалось выше, вода может передвигаться по всем трем путям (апопластным, трансмембранным, симпластным), однако доля участия того или иного из них зависит от конкретных условий, особенно от силы присасывающего действия транспирации.
В активном участии паренхимных клеток при проведении воды по корню можно убедиться в опытах с «рукавичками» и «воздушными» корнями.
Модельные системы для изучения природы корневого давления и его метаболической составляющей. В лаборатории профессора Жолкевича В.Н. (ИФР РАН) были разработаны две модельные системы для изучения «природы корневого давления – «рукавички» и «воздушные» корни.
Опыты с «рукавичками». «Рукавичка» - это апикальная часть зародышевого корня 5-7 дневного этиолированного проростка кукурузы, лишенная центрального цилиндра, при извлечении которого разрыв происходит по клеткам эндодермы (рис. 10). Следовательно, «рукавичка» не имеет ни ксилемы, ни, тем более, ксилемного сока, то есть исключается осмотическая составляющая корневого давления. Если рукавичку закрепить вертикально в воде так, чтобы верхняя часть находилась над водой, то во внутреннюю полость рукавички начинает поступать жидкость, которая нагнетается вверх, то есть наблюдается экссудация. Она возникает несмотря на полное отсутствие в полости рукавички какого-либо раствора, то есть в условиях, исключающих возможность функционирования рукавички подобно осмометру. На самом раннем этапе заполнение рукавички экссудатом осуществляется за счет метаболической составляющей. По мере заполнения полости рукавички экссудатом появляется и включается в транспортные потоки осмотическая составляющая.
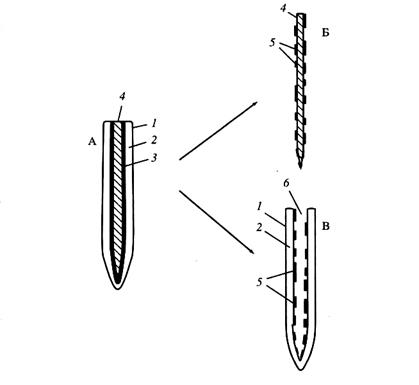 |
Рис.3. Схема получения «рукавички» из отделенного корня Zea mays L.: А – апикальная часть корня, Б – удаленный центральный цилиндр, В – «рукавичка». 1 – ризодерма, 2 – кора, 3 – эндодерма, 4 – центральный цилиндр, 5 – остатки клеток разорванной эндодермы, 6 – образовавшаяся после удаления центрального цилиндра продолговатая пустая полость.
Было выявлено, что скорость экссудации у рукавичек более чем в два раза превышает экссудацию целых корней (с центральным цилиндром). По данным Л.В. Можаевой, Н.В. Пильщиковой и Н.В. Зайцевой, найденное различие связано с повышенной сократительной способностью коры, состоящей в основном из паренхимных клеток. В центральном цилиндре доля паренхимных клеток мала. Таким образом, сокращаются, по-видимому, именно паренхимные клетки или их компоненты.
Опыты с метаболическими ингибиторами и стимуляторами показали, что экссудация рукавичек значительно чувствительнее к их действию, чем экссудация целых корней. Ингибиторы (2,4-динитрофенол и олигомицин – разобщитель окисления и фосфорилирования; КСN, ротенон, антимицин А – ингибиторы звеньев дыхательной цепи; циклогексимид – ингибитор синтеза балка и дыхания; колхицин, цитохалазин – блокаторы контрактильных систем; оуабаин – ингибитор строфантинчувствительной АТФазы) подавляли экссудацию рукавичек значительно сильнее, чем целых корней. Метаболические стимуляторы, напротив, увеличивали экссудация рукавичек значительнее, чем целых корней. В качестве стимуляторов использовали: ИУК, кинитин, гибберелловую кислоту, CaCl2, нейромедиатор ацетилхолин, аденин и др.
В опытах с ингибиторами возрастало осмотическое давление экссудата. Это происходило за счет того, что ингибиторы ослабляли способность паренхимных клеток корня удерживать растворенные вещества, которые усиленно выделяются в сосуды ксилемы. Одновременно увеличивалась и гидравлическая проводимость за счет повреждения мембран под действием ингибиторов.
Таким образом, приведенные опыты с рукавичками выявили чувствительность экссудации к метаболическим регуляторам с различным механизмом действия, что свидетельствует о наличии сложной, многоступенчатой системы эндогенной регуляции корневого давления.
Опыты с «воздушными» корнями. В опытах В.Н. Жолкевича было показано, что к экссудации способны корни совсем не поглощающие воду извне и, стало быть, вряд ли способные функционировать подобно осмометру. Были взяты отделенные корни, их вынимали из воды и тщательно обсушивали фильтровальной бумагой, а затем сжатым воздухом. Сухие корни, их назвали «воздушными», помещали в плотно закрывавшиеся пустые сосуды. В этих условиях корни не только испаряли воду, но и выделяли экссудат. Что же заставляло их это делать, и за счет чего формировался экссудат?
У «воздушных» корней нет другой возможности формировать экссудат, кроме как за счет эндогенной жидкости, отнимаемой от собственных тканей. Паренхимные клетки сами активно выделяют жидкость в сосуды ксилемы вследствие генетически запрограммированной потребности снабжать водой побеги и благодаря своей полярности. Это выделение напоминает секрецию.
Таким образом, в данном случае метаболическая составляющая проявляется в «чистом виде», а воздушные корни представляют собой очень удобную модельную систему, при работе с которой впервые удалось разграничить поглощающую и нагнетающую деятельность корня и изучать последнюю в отсутствие поглощения воды из наружной среды, то есть исключить функционирование корня подобно осмометру. Экссудация воздушных корней осуществляется только за счет метаболической составляющей.
Экссудация воздушных корней энергозависима (тормозится 2,4-динитрофенолом), стимулируется кальцием и подавляется его антагонистом – пипольфеном. Она чувствительна к блокаторам контрактильных систем – 2,3-бутандин-моноксиму и колхицину, а также нейромедиатору – норадреналину. Чувствительность воздушных корней ко всем испытанным агентам значительно превышает чувствительность водных корней (погруженных в воду).
Температурный коэффициент Q10 интенсивности экссудации воздушных корней значительно превышает Q10 водных корней. Известно, что температурный коэффициент экссудации за счет одной осмотической составляющей близок к единице.
Участие энергозависимых контрактильных систем паренхимных клеток в формировании метаболической составляющей корневого давления. Уже подчеркивалась энергозависимость экссудации и её чувствительность к блокаторам контрактильных систем. Опыты, выполненные в лаборатории В.Н. Жолкевича с корнями 5-7-дневных проростков кукурузы (таблица 4), показали торможение экссудации по сравнению с контролем блокираторами контрактильных систем: d-тубокурарином (миорелаксантом); цитохалазином Б (дезинтегратором микрофиламентов, блокирующим работу актомиозина, но не влияющего при этом на АТФазную активность миозина);2,3-бутандион-моноксимом (ингибитором миозина, подавляющем его АТФазную активность); латрункулином Б (специфическим ингибитором полимеризации актина, то есть перехода G-актина в F-актин); колхицином (дезинтегратором микротрубочек, блокирующим работу тубулиндинеиновых белков).
Таблица 4. Влияние блокаторов контрактильных систем и дыхательного энергообмена по отдельности и при совместном применении в различных сочетаниях на экссудацию отдельных корней 5-7-дневных проростков Zea mays L.
| Вариант | Подавление экссудации, % от контроля | |
| полученное экспериментально | рассчитанное как сумма отдельных эффектов | |
| ЦХБ, 1 • 10-6 М БДМ, 1 • 1О-3 М ЛАТБ, 1 • 10-6 М КХЦ, 1 • 10-3 М ДНФ, 2,5 • 10-4 М КЦХФГ, 1 • 10-6М ОМ, 10 мг • л -1 ЦХБ, 1 • 10-6 М + БДМ, 1 • 10-3 М ЦХБ, 1 • 10-6 М + ЛАТБ, 1 • 10-6 М ЦХБ, 1 • 10-6 М + ДНФ, 2,5 • 10-4 М ЦХБ, 1 • 10-6 М + КЦХФГ, 1 • 10-6 М БДМ, 1 • 10-3 М + ЛАТБ, 1 • 10-6 М БДМ, 1 • 10-3 М + ДНФ, 2,5 • 10-4 М ЛАТБ, 1 • 10-6 М + КХЦ, 1 • 10-3 М ЛАТБ, 1 • 10-6 М + ДНФ, 2,5 • 10-4 М ЛАТБ, 1 • 10-6 М + ОМ, 10 мг • л -1 ДНФ, 2,5 • 10-4 М + КЦХФГ, 1 • 10-6 М | 34,8±3,9 23,7±3,7 32,9±2,0 27,4±2,8 33,5±4,5 30,0±2,7 35,5±3,1 40,4±2,2 58,7±3,6 53,4±4,0 45,9±1,8 47,2±2,7 53,4±3,0 58,0±2,1 56,2±3,4 57,3±1,8 56,0±4,1 | - - - - - - - 58,3 67,7 60,5 64,8 56,6 57,2 60,3 66,4 68,4 63,5 |
Торможение экссудации происходило за счет агентов, имеющих различный механизм действия, но вызывающих сходный результирующий эффект: разобщителей окисления с фосфорилированием (2,4-динитрофенол, карбонилцианид-3-хлорфенилгидразон); ингибиторов окислительного фосфорилирования (олигомицин), нарушающих энергетический обмен при дыхании и тем самым препятствующих энергообеспечению всех энергозависимых жизненных функций, в том числе и механической работе контрактильных белков.
О специфическом действии перечисленных ингибиторов свидетельствует аддитивность их действия при совместном применении в разных сочетаниях. Аддитивность указывает на существование различных мишеней, на которые нацелено действие названных ингибиторов.
В опытах с отделенными корнями, помимо ингибиторов контрактильных систем, испытано влияние их антагонистов – стабилизаторов этих систем, а именно стабилизатора F-актина-фаллоидина и стабилизатора микротрубочек – таксола. Оба агента значительно стимулировали экссудацию. Их действие оказалось противоположным действию латрункулина Б и колхицина, причем наблюдалась практически полная аддитивность их стимулирующих эффектов.
Автоколебательный характер нагнетающей и водопоглощающей деятельности корня. Двухфазность процессов водообмена корня и роль энергозависимых контрактильных систем. Наблюдения за кинетикой водообмена отделенных корней выявили импульсную ритмичность как экссудации, так и водопоглощения. Оба названных процесса обнаруживают автоколебания с периодом 2-3 мин. Существенно при этом, что автоколебания находятся в противофазе, а именно максимуму экссудации соответствует минимум водопоглощения и наоборот (рис. 11). Данный факт свидетельствует о том, что при движении в радиальном направлении вода не просто пассивно «притягивается» через корень по непрерывному, плавному градиенту осмотического давления между наружным раствором и ксилемным соком (как постулирует осмотическая концепция), а поступление воды в корень и её выделение при экссудации представляют собой две ритмически чередующиеся фазы водообмена. Во время первой преобладает поступление воды, во время последующей – выделение экссудата. Чередование фаз обусловлено соответствующими колебаниями тургорного давления клеток.
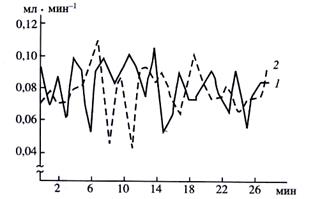 |
Рис.13. Импульсная ритмичность водообмена корня проростка Zea mays L. после резкого повышения температуры: 1 – экссудация, 2 – поглощение воды.
Впервые подобную мысль высказали Л.В. Можаева, Н.В. Пильщикова. Они предположили, что чередование максимумов выделения экссудата и поглощения воды сопровождается чередованием сокращения и расслабления корня: максимум выделения экссудата совпадает с сокращением, минимум – с расслаблением. Для проверки предположения они использовали специальный прием – резкую смену температуры в окружающей корни среде (с 15 до 30ºС). А затем измеряли поперечное сечение корня в норме и при воздействии динитрофенола или парахлормеркурибензоата (ингибитора активности контрактильных белков). В корне при разной смене температуры наступал выброс экссудата, сопровождающийся сокращением площади поперечного сечения как центрального цилиндра, так и коры, но особенно последней. Однако в присутствии ингибиторов не наблюдалось ни выброса экссудата, ни сокращения площади поперечного сечения корня. Следовательно, при ингибировании работы контрактильных белков или нарушении энергоснабжения импульсная ритмичность деятельности корня отсутствует.
Итак, короткопериодичные автоколебания экссудации и водопоглощения, особенно их асинхронность, во-первых, энергозависимы, а, во-вторых, зависят от работы контрактильных систем – как микрофиламентов, так и микротрубочек. За счет функционирования этих систем паренхимные клетки (симпласт) пульсируют, периодически сокращаясь и расслабляясь. Подобные изменения возможны благодаря эластической растяжимости клеточных стенок. Иными словами, контрактильные системы индуцируют ритмические микроколебания потенциала давления (гидростатического или тургорного давления), внешне проявляющиеся в ритмически чередующихся короткопериодных автоколебаниях экссудаций и водопоглощения. Именно таким путем формируется метаболическая составляющая корневого давления. Контрактильные белки могут выступать, например, в качестве сфинктеров, регулирующих просветы водных каналов в плазмодесмах или диаметрэндоплазматических трубок симпласта, тем более, что эти белки распределены вдоль всей мембранной системы эндоплазматической сети. Весьма существенно и значение цитоскелета в создании клеточной полярности, без которой возникновение и поддержание однонаправленного водного тока едва ли возможно.
Математическое моделирование позволило сделать вывод, что при импульсном характере переноса может быть перемещено значительно больше вещества, чем при равномерном переносе. Вероятно, именно поэтому короткопериодные автоколебания свойственны многим биологическим процессам.
Физиологическая полярность как необходимый фактор возникновения и поддержания однонаправленного водного тока при функционировании метаболической составляющей корневого давления. Возникновение однонаправленного водного тока во время фазы сокращения возможно лишь при полярности свойств отдельных клеток или же всего симпласта (симпластического домена) в целом, в частности, при полярности гидравлической проводимости.
Д.А. Сабинин подчеркивал необходимость различий в характере обмена веществ при транспорте воды в корне.
Л.В. Фоминым было обнаружено существование различия эластической растяжимости клеточной стенки в противоположных концах клетки. Благодаря этому создается полярная сократимость стенки в противоположных частях клетки, что приводит к различию тургорного и осмотического давлений. Это является предпосылкой возникновения однонаправленного движения воды.
В опытах В.Н. Жолкевича с мукором Pilobolus umbonatus Buller обнаружена взаимосвязь однонаправленного водного тока и электрической поляризации клетки (рис. 12). Гриб поглощает воду гифами, погруженными во влажный субстрат, а через спорангиеносец, представляющий гигантскую клетку, выделяют её в виде обильной гуттации. Микроэлектродные измерения выявили наличие четко выраженного апикально-базального градиента мембранного потенциала. Оказалось, что вода передвигается в направлении возрастания потенциала, а интенсивность гуттации положительно коррелирует с величиной названного градиента. При воздействии анестетиков (паров серного эфира или хлороформа) одновременно исчезают и гуттация, и градиент мембранного потенциала.
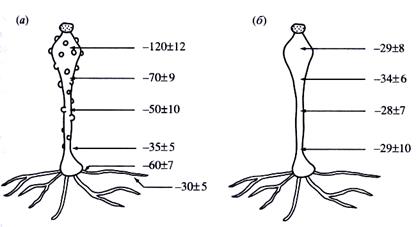 |
Рис. 12. Взаимосвязь между гуттацией и градиентом мембранного потенциала (в мВ) у гигантской клетки Pilobolus umbonatus Buller: а – контроль, б – при воздействии серного эфира или хлороформа (10 мин).
Взаимодействие цитоскелета, нейромедиаторов, фитогормонов и кальция при функционировании метаболической составляющей корневого давления. Автоволновая природа транспорта воды в корне естественно требует необходимости взаимодействия соответствующих регуляторных и сигнальных систем. Что же представляют собой эти системы? Они включают: контрактильные системы (элементы цитоскелета), нейромедиаторы, фитогормоны, кальций. Участие контрактильных систем, фитогормонов и кальция в регуляции транспорта воды представляется несомненным. Однако предположение об участии нейромедиаторов нуждается в мотивировке.
В середине 1970-х годов были начаты работы (в лаборатории В.Н. Жолкевича) с нейромедиаторами ацетилхолином и d-тубокурарином. Первый - медиатор нервного возбуждения, а второй – его антагонист, блокирующий передачу возбуждения с двигательного нерва на мышцу. Многие считали, что данные агенты не имеют прямого отношения к физиологии растений. Однако постепенно выяснилось, что подвижность и хеморецепция не являются привилегией животных, а имеют всеобщее распространение среди живых организмов. Разница заключается лишь в том, что у животных эти свойства выражены сильнее, а двигательная и рецепторная системы более дифференцированы и совершенные. Поэтому они раньше привлекли к себе исследователей. Выделенный из мышечных тканей белок актомиозин затем был обнаружен в цитоскелете (в контрактильной системе) клеток. Если в немышечных (растительных) клетках функционируют контрактильные белки, то должен существовать механизм, регулирующий их работу. Опыты с отделенными корнями проростков кукурузы показали, что ингибиторы контрактильных белков тормозили экссудацию, а нейромедиаторы (ацетилхолин, адреналин, норадреналин, серотонин), фитогормоны (ИУК, зеатин, гибберелловая кислота) и кальций неизменно стимулировали её. В присутствии ингибиторов контрактильных белков или энергообмена при дыхании (ДНФ, КЦХФГ) стимулирующий эффект всех нейромедиаторов и фитогормонов полностью снимался (как будто в инкубационной среде находились одни только ингибиторы). При совместном применении нейромедиаторов и фитогормонов в различных сочетаниях не отмечено аддитивности их стимулирующего эффекта на экссудацию.
Таким образом, стимулирующее воздействие на нагнетающую деятельность корня как нейромедиаторов, так и фитогормонов невозможно без функционирования белков цитоскелета и/или при нарушенном энергоснабжении. Это обстоятельство может свидетельствовать: 10 о ведущей роли белков цитоскелета в создании корневого давления; 2) о том, что стимулирующее действие нейромедиаторов и фитогормонов нацелено прежде всего на энергозависимые контрактильные системы. Вероятно поэтому не наблюдается аддитивность стимуляции экссудации при совместном применении нейромедиаторов и/или фитогормонов. Центральное звено, без которого невозможно действие этих стимуляторов, одно и то же – функционирование цитоскелета. Полагают, что цитоскелет обеспечивает восприятие и передачу сигналов.
Полное раскрытие всех механизмов, посредством которых паренхимные клетки направляют и регулируют ток воды, пока остаются делом будущего.








