 2015-07-14
2015-07-14 1144
1144Как уже указывалось, насекомоядные являются прямыми филогенетическими предшественниками грызунов, хищных и приматов.
Исследование конструкции ассоциативных систем мозга у крыс, кроликов, кошек, собак и низших обезьян позволили прийти к следующему заключению.
Первый уровень — грызуны, мозг которых близок к насекомоядным. У грызунов отсутствует четкая дифференциация коры и таламуса на специфические и ассоциативные зоны. Известная диффузность представительства сенсорных систем в больших полушариях головного мозга грызунов коррелирует со сравнительно низким уровнем их аналитико-синтетической деятельности. Слабая выраженность морфологической дифференциации и функциональной специализации полисенсорных структур у грызунов является фактором, определяющим несовершенство интегративной функции мозга.
У белых крыс формирование условного рефлекса на зрительно-слуховой комплекс оказалось также безрезультатным, неоднократные тренировки этой трудной задачи приводили к невротическим срывам. В то же время зрительно-тактильный комплекс выработать удалось, хотя он характеризовался нестабильностью. Примерно аналогичная картина выявилась у кроликов, у которых дифференцирование светового и звукового компонентов
от их одновременного предъявления в комплексе не достигало выше 50—60% -го уровня. Это отражает низкий уровень аналитико-синтетической деятельности насекомоядных и грызунов.
Второй уровень — хищные, у которых впервые появляются в коре больших полушарий развитые лобные и теменные ассоциативные поля и соответствующие структуры таламуса. Для хищных характерны существенные структурные и функциональные различия ассоциативных таламокортикальных систем от других мозговых структур. Это — наличие сложных нейроносинаптичес-ких компонентов, к которым конвергируют сенсорные потоки, несущие биологически значимую информацию. Причем таламопариетальная система отражает усложнение актов пространственной ориентировки и формирование механизмов, составляющих текущий сенсорный фон для выполнения целенаправленных поведенческих актов. Таламопариетальная система — следствие усложнения конструкции и связей зрительной сенсорной системы, она обеспечивает первичной межсенсорный синтез, формирует сложный комплекс обстановочной афферентации и систему «схемы тела».
Таламофронтальная система включается в корковый отдел соматической сенсорной системы с одновременной проекцией на нее лимбических аппаратов мозга. Таламофронтальная система участвует в организации предпусковой интеграции и программировании сложных поведенческих актов. Здесь в большей мере отражены процессы мотивационно-эмоциональной окраски поведенческих актов благодаря прямым связям этой системы со всем лимбическим комплексом. В пределах отряда хищных лобные отделы неокортекса усложняются, увеличиваются их размеры и функциональная роль в организации сложных форм поведения, требующих мобилизации механизмов краткосрочной и долгосрочной памяти.
У хищных (кошки, собаки) выработка условного рефлекса на разномодальный комплекс выполняется за 10— 15 опытов. Уровень дифференцирования светового и звукового компонентов от комплекса достигает 70—90%, но для этого требуется регулярная тренировка. Иными словами, сигнальное значение комплекса поддерживается
при условии угашения реакций на отдельные компоненты. Следовательно, у хищных существует более высокий уровень аналитико-синтетическои деятельности, заключающейся в способности к интеграции разномо-дальных сигналов в целостный образ.
Другой важной стороной интегративной функции мозга является степень развития процессов памяти и основанное на них свойство прогнозирования предстоящего поведения. В естественных же условиях существования животных любая поведенческая адаптация относительна и имеет вероятностный характер. Поэтому применительно к биологическим условиям обитания животных в вероятностно изменчивой внешней среде имеет место и вероятностное прогнозирование, а значит, адаптивность поведенческих программ определяется степенью их избыточности и подвижности.
Изучалось поведение в стационарных случайных средах. На звуковой сигнал животное должно было направляться к одной из двух кормушек, причем чаще всего к той, где наиболее вероятно получало подкрепление. Степень вероятности подкрепления из каждой кормушки менялась (А. С. Батуев, И. В. Малюкова, 1979; А. И. Ка-рамян, И. В. Малюкова, 1987). Кошки способны к формированию поведения в сторону той кормушки, вероятность пищевого подкрепления из которой наиболее высока. Причем поведение их меняется в соответствии с изменением вероятности подкрепления. Повреждение фронтальных отделов неокортекса (в основном сенсомо-торной коры) сохраняет элементарные условные рефлексы, но разрушает способность к вероятностному прогнозированию.
Процессы межсенсорной интеграции развиты у хищных в достаточной мере, хотя кортикальный уровень ассоциативных систем мозга имеет перекрытие с зонами выхода эфферентных кортикальных трактов. С другой стороны, у хищных достаточно велика роль докорко-вого уровня межсенсорной интеграции и кора получает наряду с модально специфичными афферентными залпами уже переработанную неспецифическую импульсацию из ассоциативных ядер. Наконец, каждая из ассоциативных систем у хищных характеризуется доминиро-
ванием того или иного сенсорного входа, что, естественно, не способствует достижению полной гетеросенсор-ной интеграции.
Третий уровень — приматы, у которых ассоциативные структуры таламуса с их обширной и дифференцированной проекцией в лобные и теменные области коры включаются в состав самостоятельной целостной инте-гративной системы больших полушарий. Существенной особенностью приматов являются развитые кортико-кор-тикальные связи, с помощью которых ассоциативные поля могут объединяться в целостную иерархически построенную систему. Благодаря компактной системе мие-линизированных ассоциативных волокон возрастает роль кортикального уровня взаимодействия специфических сенсорных зон с ассоциативным неокортексом. Ассоциативные поля неокортекса характеризуются тонкой дифференциацией с формированием из нейронных элементов целостных структурно-функциональных ансамблей.
Спецификой ассоциативного неокортекса является конвергирование в нем множества сенсорных сообщений о биологической значимости внешней сигнализации в соответствии с доминирующей мотивацией по независимым друг от друга афферентным каналам. При этом возрастает роль собственно кортикального уровня межсенсорной интеграции и обеспечения процессов краткосрочной памяти. После разрушения либо лобных, либо теменных ассоциативных полей коры не только глубоко страдают процессы межсенсорного синтеза, ориентировочно-исследовательская деятельность, краткосрочная память, но и формирование более простых форм услов-норефлекторной деятельности.
Функциональная значимость отдельных ассоциативных систем расширяется и уточняется в сравнении с хищными. Утрачивается преобладание какого-либо одного сенсорного входа, а следовательно, расширяются возможности их интеграции. Возникает топографическая разнесенность ассоциативных полей от собственно афферентных корковых формаций, что снижает удельное значение сенсомоторной интеграции и расширяет роль коры в осуществлении межсенсорного афферентного синтеза. Возникает все большая их взаимозависимость
для обеспечения деятельности целостной интегративнои системы полушарий.
Для приматов (низшие обезьяны) выработка условных рефлексов на одновременный комплекс представляет собой относительно легкую задачу, ибо в процессе применения комплексного раздражителя компоненты самопроизвольно утрачивают сигнальное значение и сформированные условнорефлекторные связи сохраняются месяцами без дополнительной тренировки. У обезьян можно выработать условный рефлекс даже на трехчленный комплекс из разномодальных сигналов. Это свидетельствует о более высоком уровне аналитико-син-тетической деятельности мозга обезьян в сравнении с хищными, что находит отражение и в большей структурной дифференциации и функциональной специализации ассоциативных таламокортикальных систем.
Обезьяны легко справляются с задачами на вероятностное прогнозирование, однако после разрушения в области ассоциативной лобной коры лишаются этой способности, их поведение приобретает однообразный пер-северативный характер.
Очевидно, способность использования предыдущего опыта, записанного в долгосрочной памяти, для прогнозирования поведения в стационарных случайных средах, претерпевает существенные эволюционные преобразования, которые определяются степенью развития интегра-тивных систем мозга, уровни дифференциации которых коррелируют со степенью совершенства аналитико-син-тетической деятельности и организации сложных форм поведения.
ОНТОГЕНЕЗ АССОЦИАТИВНЫХ СИСТЕМ МОЗГА
Изучение динамики формирования ассоциативных систем мозга показало гетерохронность этого процесса, что, по-видимому, обусловлено включением отдельных звеньев таламокортикальных систем в обеспечение разных поведенческих актов, последовательность созревания которых определяется их необходимостью для осуществления жизненно важных функций новорожденного животного (зрело- и незрелорождающегося).
Согласно концепции системогенеза (П. К. Анохин, 1968) неравномерность созревания нервных элементов и связей между ними объясняется их вовлечением в структуру различных функциональных систем. Гетерохрон-ность их созревания определяется значимостью для выживания организма особенно в критические периоды жизни (см. гл. 4), когда новорожденное животное вступает в непосредственное соприкосновение с окружающей средой. Анатомически и функционально первыми созревают те сенсорные механизмы (соматические, акустические), которые обеспечивают выживание на начальных этапах индивидуального развития.
В ассоциативных ядрах таламуса обнаружены проекции в кору больших полушарий уже у новорожденных котят (В. П. Бабминдра, Л. А. Васильева, 1987). В то же время неспецифические таламические ядра формируют свои проекции на кору несколько позже, их основная роль состоит в контроле внутриталамической активности.
Несмотря на раннее формирование ассоциативных систем, их окончательное созревание происходит в течение довольно длительного периода и заканчивается у кошек между 2-м и 3-м месяцами жизни. Именно к этому времени у котят впервые формируется полноценный условный рефлекс на одновременный комплекс, приближающийся по своим характеристикам к аналогичному условному рефлексу взрослого животного (Л. А. Васильева, Л. В. Черенкова, 1986).
Когда первый критический этап постнатального развития преодолен, наступает период бурной дифференциации мозговых структур и возникновения между ними двусторонних связей диффузного характера. Формируется та структурно-функциональная матрица, которая послужит основой для дальнейшего развертывания процессов координации и выделения локальных функциональных структур.
Таким образом, принцип развития — от диффузного неспецифического к локальному специфическому (А. И. Карамян, 1976) — общебиологическая закономерность, которой подчиняется и динамика развития ассоциативных систем мозга.
Наконец, третий этап связан с моментом формирования тормозных координационных механизмов как в самой коре, так и в глубоких структурах. Появление таких механизмов обеспечивает тонкую специализацию как сенсорных, так и ассоциативных систем мозга, а значит, и различных целостных поведенческих актов.
§ 44. АССОЦИАТИВНЫЕ
СИСТЕМЫ И СЕНСОРНАЯ ФУНКЦИЯ МОЗГА
Прежде чем произойдет какая-нибудь приспособительная реакция, осуществляется процесс восприятия сигналов. Но живой организм заинтересован не в самих воздействиях как таковых, а в том, о чем они сигнализируют и соответственно не в оценке их физических параметров, а в тех соотношениях, которые с их помощью передаются. Вместе с тем понятно, что отнесение сигнала к тому или иному классу, то есть его распознавание, неизбежно основано на выделении ряда физических характеристик, образующих область признаков данного класса сигналов.
Из сказанного выше следует, что сенсорная функция мозга заключается в определении сигнальной (биологической) значимости сенсорных стимулов на основе анализа их физических характеристик. Под биологической значимостью понимается направленность реакций живого организма, которая определяется его доминирующей мотивацией, возникающей при отклонении параметров устойчивого неравновесия, а также информацией, извлекаемой из окружающей среды и прошлого жизненного опыта. Для оценки биологической значимости сенсорных сигналов анализ их физических характеристик является необходимой, но недостаточной операцией. Биологическая значимость сигнала реализуется в активации некоей совокупности эффектор-ных аппаратов. Следовательно, извлечение биологически полезной информации с целью ее использования для формирования двигательных актов основано на преобразовании входной кодовой комбинации активности нервных элементов сенсорных систем в реакцию испол-
нительных аппаратов, что по своей сути представляет процесс декодирования сенсорных сообщений. Тогда оценка биологической значимости сигнала сводится к установлению соответствующей закономерности взаимодействия нервных элементов сенсорных и двигательных систем мозга.
Выход на эффекторные элементы должен обладать свойстзом избирательности, чтобы избавить систему от неадекватных реакций. Выделение экологически значимых признаков (обычно достаточно простых) у животных низких уровней эволюционного развития может осуществляться в самих сенсорных системах (иногда уже на рецепторном уровне) вследствие жесткой организации нервных связей. Однако у более высокоорганизованных животных (хищных, приматов) эта операция уже не может ограничиться специфическими образованиями сенсорных систем. Ведь по мере развития уровня организации животных рецептуар стимулов, имеющих сигнальное значение, как правило, расширяется. Поэтому возрастание активности организма как системы должно быть связано с возможностью перехода избирательного приема от одних сигналов к другим и в соответствии с этим — с появлением новых внутримозго-вых конструкций.
Нервные образования, входящие в систему оценки биологической значимости сигналов, должны удовлетворить ряду требований. Во-первых, они должны быть связаны с различными сенсорными системами. Во-вторых, они должны быть связаны с мотивациогенными структурами лимбической системы, ибо значимость тех или иных сигналов определяется на основе доминирующей мотивации. В-третьих, учитывая, что значимость одного и того же сенсорного стимула зависит от всей окружающей ситуации, то есть от показаний других сенсорных систем, следует допустить наличие конвергенции полимодальной импульсации и способность к пластическим перестройкам активности нейронов. Наконец, в-четвертых, эта система должна быть связана с регуляцией целостных двигательных актов на основе избирательного характера реагирования. Изложенным требованиям удовлетворяют таламокортикальные
ассоциативные системы мозга: таламопариетальная и таламофронтальная.
Получены доказательства (А. М. Иваницкий, 1976) того, что ранние компоненты суммарного вызванного потенциала мозга человека отражают поступление в кору информации о физических характеристиках сенсорных сигналов, а поздние — являются отражением значимости сигнала для организма. У больных с психическими расстройствами, имеющих определенный дефицит одного из видов информации, возникают стойкие изменения соответствующих фаз вызванных потенциалов. В особенности выделяются ассоциативные области неокортекса в связи с выраженной конвергенцией к ним разномодаль-ной импульсации как основы оценки сигнальной значимости стимула в контексте соответствующих воздействий.
Именно этот механизм у высших позвоночных способствует селективному отбору одних видов информации с одновременным сопряженным торможением других сенсорных влияний. При этом у животных доминирующая мотивация возникает из биологической потребности в результате сдвигов гомеостаза. Однако доминирующая мотивация формируется при воздействии стимулов, ранее связанных с определенными подкреплениями. Живая система способна предвидеть изменения гомеостаза с помощью сигналов, предвосхищающих сдвиг параметров от нормы или возвращение к ней. В связи с этим другие сенсорные стимулы способны изменять значимость сигнала, если они меняют доминирующую мотивацию. И тогда значимый стимул является сигналом, реагирование на который сопряжено с большей вероятностью достижения желаемого результата в соответствии с доминирующей мотивацией и прошлым жизненным опытом. Наличие сопряженных признаков в комплексном сенсорном воздействии позволяет уменьшить неопределенность выбора поведенческих реакций, необходимых для получения требуемого результата.
Роль ассоциативных систем в оценке биологической значимости сенсорных сигналов отчетливее видна по отношению к зрению и слуху. Конечно, сигнальное значение имеют все сенсорные стимулы, но способностью опережающего воздействия обладают в первую очередь
зрительные, слуховые, а также обонятельные сигналы. Особенностью структурной организации телецептивных входов в ассоциативные системы является множественность путей проведения к ним слуховых и зрительных афферентаций.
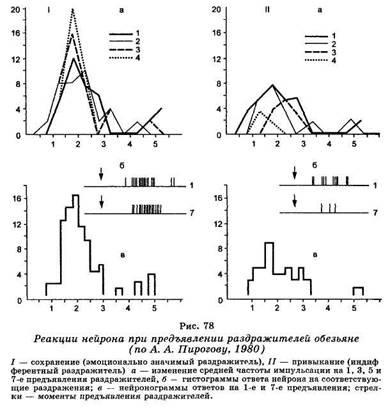
Для целей определения биологической значимости механизмы конвергенции обеспечивают полифункциональность одного и того же сигнала, то есть зависимость его смысла от ситуации, в которой он возникает. Степень значимости любого агента внешней среды зависит не только от определенной мотивации, но и от сопоставления этого агента с другими факторами среды, когда он выступает в качестве компонента более сложного сенсорного комплекса. Именно поэтому наиболее полная сигнальная значимость фактора окружающего пространства может реализовываться с участием тех структур мозга, куда приходит информация о разных по сенсорным качествам факторах среды, то есть по механизму одновременного гетеросенсорного сопоставления. Одним из проявлений динамического характера конвергенции в ассоциативные зоны коры является свойство привыкания их электрических реакций при использовании однообразной ритмической стимуляции. Степень развития привыкания находится в прямой зависимости от биологической значимости применяемого сигнала. Например, стабильные ответы нейронов лобных долей у бодрствующих обезьян возникают лишь в том случае, если раздражители прямо или косвенно связаны с эмо-ционально-мотивационной сферой животного, необычны для него или сигнализируют об опасности, то есть сопровождаются четкой ориентировочной реакцией. Привыкание ответов наблюдается в тех случаях, когда используемые раздражители являются индифферентными для животного или хорошо знакомы ему (рис. 78). Если же эти агенты сопровождаются биологически значимым подкреплением по правилам выработки условного рефлекса, то они приобретают свойства условного сигнала с одновременной стабилизацией реакций нейронов лобной коры в ответ на их применение.
Как привыкание, так и сенситизация отражают дистрибутивный характер данных пластических реакций
мозга и совпадают с закономерностями проявления ориентировочного рефлекса. Поэтому предполагается, что «новизна» является тем информативным признаком сенсорных стимулов, с которого начинается их оценка в ассоциативных системах мозга. Привыкание к повторным стимулам, вероятно, связано с процессами формирования следов кратковременной памяти (Е. Н. Соколов, 1981).
С помощью ориентировочной реакции устанавливаются соответствующие воспринимающие поверхности для наилучшего восприятия внешнего раздражения. При размещении источника телецептивных стимулов по средней линии головы наблюдается их наиболее высокое пространственное разрешение. Избирательность к этой фи-
зиологически значимой области пространства применительно к акустическим сигналам обнаружена для нейронов мозжечка и сенсомоторной ассоциативной коры (Я. А. Альтман, 1972; Г. А. Куликов, 1989). Участие фронтальной и теменной областей коры в обеспечении точности ориентировочных реакций установлено и для зрительных сигналов, по отношению которых отмечена и пространственная, и дирекциональная избирательная чувствительность соответствующих нейронов. Более того, после повреждения теменной ассоциативной области отчетливо выступает нарушение пространственного восприятия зрительных стимулов и адекватных двигательных реакций, то есть нарушения пространственного согласования зрительно-моторной координации.
Система, оценивающая биологическую значимость сенсорных стимулов, должна обладать избирательным характером реагирования на их отдельные значимые признаки и/или совокупность последних. Поэтому логичен вопрос о том, как отражаются в реакциях ассоциативных структур мозга различные физические параметры сенсорных сигналов.
Известно, что нейроны фронтальной ассоциативной коры принимают зрительную импульсацию, не связанную с поточечным описанием контура зрительных сигналов, но отражающую их временные и пространственные параметры. Кроме того, показано, что нейроны фронтальной коры обладают свойством избирательного реагирования на биологически значимые признаки коммуникативных акустических сигналов. Существенно то, что для млекопитающих (в отличие от низших позвоночных) лишь небольшое число сигналов может иметь четкое «ключевое» значение, которое в большей мере определяется доминирующим мотивационным фоном для данного животного. Поэтому становится понятным участие в оценке биологической значимости сигнала тала-мофронтальной ассоциативной системы, тесно связанной с лимбическими структурами мозга, имеющими прямое отношение к формированию мотивационных состояний (Г. А. Куликов, 1989).
В связи с этим закономерен вопрос, как отражается в характере электрических реакций фронтальной коры
процесс приобретения ранее индифферентным звуковым стимулом определенного сигнального значения при выработке условного рефлекса. Специальные исследования показали, что преобразование ранее неэффективного звукового тона в мотивационно окрашенный сигнал оборонительной реакции животного сопровождается возникновением избирательного реагирования элементов фронтальной коры именно на данную частоту заполнения тонального сигнала. Однако по мере стабилизации и закрепления условного рефлекса такая избирательность выражалась слабее и, наконец, исчезала, что свидетельствует о динамической роли фронтального неокортекса в формировании условнорефлекторной сенсомоторной интеграции.
Описанные выше закономерности избирательного реагирования на сигнально значимые агенты среды могут служить физиологическими предпосылками для трактовки механизмов сенсорного внимания (Н. Ф. Суворов, О. П. Таиров, 1986).
В самом общем смысле внимание рассматривается как один из механизмов устранения избыточности сенсорных сообщений, который участвует как в избирательном регулировании сенсорных сообщений, так и фильтрации информации, извлекаемой из систем кратковременной и долговременной памяти. Принято думать, что процессы фильтрации осуществляются уже в специфических образованиях сенсорных систем. Между тем в ряде работ мнение об уменьшении избыточности информации основано на данных об уменьшении плотности потока импульсации при переходе от нижележащих уровней сенсорных систем к вышележащим и на постулировании использования в их деятельности двоичного кода. Этот вопрос, однако, остается не решенным с нейрофизиологических позиций, и проблемой «внимания» преимущественно владеют психологи и психофизиологи.
Сам А. А. Ухтомский (1923), сформулировав принцип доминанты как физиологическую основу акта внимания, писал, что из множества действующих рецепций доминанта вылавливает группу рецепций, которая для нее в особенности биологически значима.
Определение вектора поведения (по принципу доминанты) основано на взаимодействии биологически значимых экстероцептивных (окружающая среда) и инте-роцептивных (мотивационных) воздействий, основным субстратом установления которого являются таламокор-тикальные ассоциативные системы мозга.
Следовательно, целостная сенсорная функция мозга обеспечивается содружественной деятельностью сенсорных и ассоциативных систем и направлена на организацию адаптивных движений и действий. Последнее лежит в основе согласования движения с чувствованием (И. М. Сеченов, 1863).
§ 45. АССОЦИАТИВНЫЕ СИСТЕМЫ МОЗГА И ПРОГРАММИРОВАНИЕ ПОВЕДЕНИЯ
Одним из основных атрибутов любого сложного целенаправленного движения является формирование предварительных программ.
Роль программы в структуре двигательного акта должна рассматриваться с учетом биологической мотивированности движения, его временных параметров, моторной дифференцированности, степени сложности координационного состава и уровня его автоматизированное™.
Стратегия и тактика движения. Биологическая мотивированность двигательного акта является основным побуждающим (инициальным) фактором для его реализации. Именно мотивации формируют целенаправленные движения, а следовательно, определяют их общую стратегию. Значит, если в основе стратегии движения находится биологическая (или социальная) мотивация, то каждый конкретный двигательный акт будет рассматриваться как шаг к удовлетворению этой мотивации, то есть будет решать какую-то промежуточную задачу или цель (рис. 79). Биологические мотивации могут приводить либо к запуску «запаянных», то есть жестких, программ, устанавливать их комбинаторику, с чем мы встречаемся у беспозвоночных и низших позвоночных
и именуем инстинктами или комплексами фиксированных действий, либо приводить к формированию новых сложных программ, определяя одновременно степень их лабильности.
П. Тейтельбаум (1967) пишет, что в тех случаях, когда действие полностью является автоматическим следствием стимула, невозможно говорить о мотивации. При этом между стимулом и ответом существуют фиксированные взаимоотношения. Мотивация «ломает» эти фиксированные связи между стимулом и реакцией с помощью процесса обучения. Например, в отличие от многих инстинктивных реакций реакция нажатия на педаль может быть «отделена» от внутреннего состояния животного. Опе-рантная ситуация, сигнал, реакция, подкрепление являются полностью произвольными, не обладающими фиксированными связями друг с другом.
Возьмем для примера такое биологическое адекватное движение у кошек, как прицельный прыжок. У одной группы кошек он условнорефлекторно закреплялся при пищевом подкреплении, у другой — при электрооборонительном (А. С. Батуев, 1960). У животных экстренно выключалась зрительная рецепция. Условные прыжки при пищевом подкреплении исчезли полностью, а при электрооборонительном — восстанавливались через две недели после операции. Следовательно, одна и та
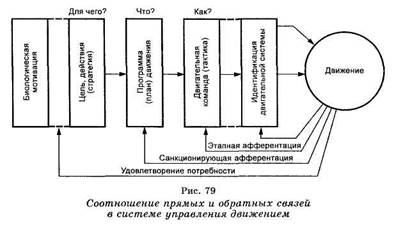
же двигательная программа (или план движения, по Прибраму) характеризовалась разной степенью устойчивости к сенсорной депривации в зависимости от биологического знака подкрепляющего агента. Образно говоря, если мотивация определяет, для чего, с какой целью осуществляется движение, то она же, определяя программу конкретного движения, обусловливает степень ее зависимости от сенсорных факторов. В качестве обратной афферентации здесь выступает удовлетворение мотивации. Поэтому цель движения, его стратегия должна быть прежде всего адаптивна по отношению к доминирующей биологической мотивации.
Программа определяет, какое конкретное движение будет осуществляться для достижения данной цели, и формирует его пространственно-временные параметры в соответствии с реальной обстановкой. Следовательно, программа движения должна быть в высокой степени адаптивной ко всем факторам окружающей среды и строиться при их одновременном и строгом учете, то есть на основе синтеза всех видов афферентации по закону муль-тисенсорной конвергенции.
В общей схеме системной организации двигательного акта существует понятие о тактике движения, которая определяет, как будет осуществляться требуемое движение, иными словами, каковы те эфферентные залпы к двигательным ядрам спинного мозга, которые определяют активацию различных мышечных групп. Последовательность включения той или иной конечности, записанная в программе, реализуется в виде последовательности импульсных залпов надсегментарных структур к различным мотонейронным пулам.
В отличие от программы двигательные команды должны быть очень адаптивными к функциональному состоянию самого скелетно-двигательного аппарата как непосредственного исполнителя этих команд. Исходя из представления о существовании настройки перед совершением движения (Я. М. Коц, 1975; Э. Эвартс, 1975), высказано предположение о том, что срочная идентификация состояния сложной системы осуществляется путем оценки реакции системы на небольшое точно дозированное возмущение (тест-импульс), которое подается
непосредственно перед или с моментом запуска команд. Причем тест-импульс подается только на те мышцы, которые должны будут участвовать в следующем движении. Реакция мышц на тест-импульс будет не настолько значительна, чтобы существенно изменить само движение, но достаточна для возбуждения мышечных веретен при изменении длины интрафузальных волокон на 5— 10 мк. Информация о реакции на тест-импульс конвергирует на некотором промежуточном уровне с основным управляющим воздействием (командой), которое корректируется в соответствии с актуальным состоянием двигательного аппарата. Учитывая временную дисперсию проведения по пирамидному тракту, легче всего допустить участие быстропроводящих его волокон (со скоростью 50-60 м/с) в реализации такого тестового воздействия и коррекции последующих нисходящих импульсных залпов уже на сегментарном уровне. Таким образом, биологически мотивированная стратегия (цель) движения, соответствующая программа конкретного двигательного акта, иерархически построенные центральные команды и система идентификации состояния мышечного аппарата представляют собой важнейшие звенья организации двигательного акта, каждое из которых корректируется соответствующими обратными связями.

