 2015-08-21
2015-08-21 974
974IE некоторых водоемах планктонные ветвисто-1ше ракообразные подобно другим планктонным нвотным совершают регулярные суточные вер-■альные миграции. В темное время суток они Ищимаются в поверхностные слои воды, а в ■мое время уходят вниз. Однако во многих Врах и водохранилищах большая часть рачков круглые сутки остается близ поверхности. I До тех пор пока условия жизни ветвистоусых Вов благоприятны, в водоемах встречаются ■иько самки, откладывающие и вынашивающие ■ выводковых камерах неоплодотворенные яйца. ■ело этих яиц различно не только у разных ви-||ов, но даже у различных особей одного вида.
Разные виды дафний образуют до 50 или даж 100 яиц на самку, босмины — до 16 j а мелкие донные виды — не больше 2 \ Яйца развиваются в выводковой Kai матери, где из них образуются сна' подвижные эмбрионы, а затем вполне сформ вавшиеся маленькие рачки, покидающие вы новую камеру и приступающие к самостоятел му существованию. Летом, при достаточном личестве пищи, рождение молоди и появл в камере новых яиц происходят каждые 1—2 ток, благодаря чему численность ветвисто;; может очень быстро увеличиваться. Обыкнов» рождение молоди сопровождается линькой cai
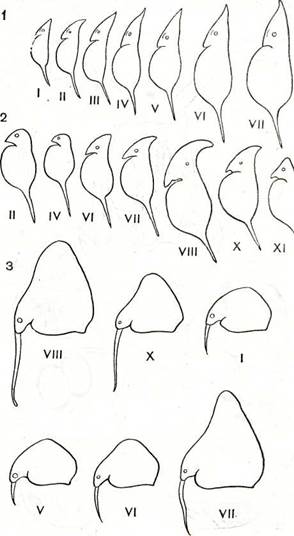
Рис. 197. Цикломорфоз планктонных ветвистоусых
1 — Daphnia cucullata; 2 — D. cristata; s — Bosmina coi (римскими цифрами обозначены месяцы, в течение которых су вует каждая форма).
У хищных ветвистоусых, например у лептодоры и битотрефеса, выводковая камера изолирована от окружающей среды и при выходе молоди каждый раз разрушается, а затем восстанавливается вновь.
Вышедшая.на свободу молодь часто линяет и быстро растет. Первые три линьки происходят через 1 —1,5 суток, а следующие через 2—3 суток. Большинство видов достигает половозрелости после третьей-четвертой линьки, т. е. через 2—6 суток после рождения. Так получаются новые поколения самок, продолжающие размножаться пар-теногенетически.
Но вот условия существования ветвистоусых рачков ухудшились вследствие понижения темпе-
Рис. 198. Размножение ветвистоусых:
I — спаривание дафний; 2 — поперечный разрез через яйцо Daph-
nia magna; s — 13 — эфиппиумы разных ветвистоусых: з — Daphnia
magna, 4 — D. balchashensis, 5 — Leydigia acanthocercoides, 6 —
Oxyurella tenuicaudis, 7 — Bosmina longirostris, s — Eurycercus la-
mellatus, 9 — Drepanothrix dentata, 10 — Ilyocriptus sordidus,
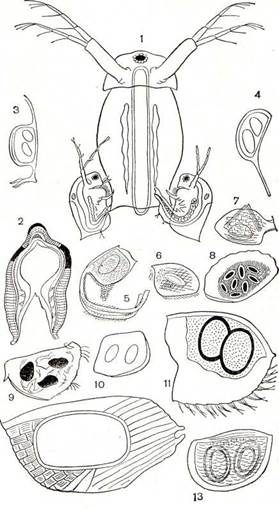 |
II — Maerothrix hirsuticornis, 12 — Alonella excisa, 13 — Moina
macrocopa.
ратуры, уменьшения количества пищи или загрязнения водоема. Это ухудшение сейчас же сказывается на судьбе яиц, находящихся в половых путях самки. Из неоплодотворенных яиц после их откладки в выводковую камеру выходят не самки, а самцы. Кроме того, часть яиц в половых путях самки подвергается второму делению созревания, в результате которого количество хромосом в яйце уменьшается вдвое. Такие яйца могут раз виваться только после оплодотворения. При слиянии яйца и сперматозоида восстанавливается полное количество хромосом.
Замечательно, что даже кратковременные отклонения от нормальных условий жизни могут прерывать процесс партеногенетического размножения. Судьба яйца определяется за 15 мин до его выхода из половой системы самки. Если в этот момент рачки подвергнутся каким-нибудь неблагоприятным воздействиям, из их яиц выведутся самцы или эти яйца будут непременно нуждаться в оплодотворении.
Самцы ветвистоусых ракообразных — карлики по сравнению с самками. Часто они в несколько раз меньше самок (рис. 198). Их передние антенны удлинены, а первая пара грудных ножек снабжена коготками, помогающими удерживать самку,
Обыкновенно самцы появляются в водоемах осенью, после значительного понижения температуры. Они прикрепляются к самкам при помощи коготков передних ног и при помощи передних антенн, причем у дафний нередко к одной самке прикрепляются два самца. Сперма проникает в выводковую камеру близ отверстий яйцеводов, а потом и в них.
Оплодотворенные яйца во многих отношения! отличаются от неоплодотворенных. Их гораздо меньше (редко больше двух), но зато они значительно богаче желтком и поэтому крупнее, чем партеногенетические. Вокруг них развиваются защитные оболочки. У Sida, Diaphanosoma, Но-lopedium и хищных видов оплодотворенные яйца снабжены двумя оболочками — плотной внутренней и клейкой наружной. Такие яйца откладываются в воду и опускаются на дно.
У большинства ветвистоусых оплодотворенные яйца откладываются в выводковую камеру, а в образовании их защитных оболочек принимает участие материнская раковина. Ее спинной край становится толще, под раковиной собираются клетки, плотным слоем окружающие яйцо или яй-1 ца. У дафний поверх этих клеток выделяется кутикула и в выводковой камере возникает так называемое седлышко, или эфиппиум, содержащее обычно два яйца. Формирование эфиппиума продолжается 2—3 суток, и самка носит его в течение 1—2 суток. Затем она линяет, и эфиппиум оказывается на свободе. Благодаря присутствию воздухоносного слоя эфиппиумы дафний плавают на поверхности воды. У некоторых других ветвис-
иоусых наружной оболочкой эфиппиума служит Вся раковина или значительная ее часть, сброшенная при линьке (рис. 198). Главная биологическая особенность оплодот-юренных яиц заключается в том, что они могут увиваться только 1госле более или менее про-влжительного периода покоя и поэтому назы-вются покоящимися. Такие яйца помогают рач-Ьм переносить всевозможные неблагоприятные ыовия; они вмерзают в лед, высыхают, сохраняя (течение длительного времени жизнеспособность. же время они служат средством расселения |етвистоусых, легко переносясь из водоема в во-рем ветром, вместе силом, присохшим к лапкам (одошавающих птиц, и т. д. Благодаря покоя-шся яйцам ветвистоусые широко распространю во всех стоячих водоемах нашей планеты. Многие виды зиму проводят в стадии покояще-р яйца, которое начинает развитие весной и 1т начало самке, приступающей к партеногене-иескому размножению. У всех ветвистоусых, I исключением лептодоры, из яйца выходит илне сформировавшийся рачок. Лептодора — действенный представитель ветвистоусых, имею- щ личинку; она вылупляется из покоящегося ща на стадии метанауплиуса (табл. 48, 17). Число известных видов ветвистоусых — около!0. Некоторые из них распространены очень шило и встречаются в самых разнообразных водое-р, другие приурочены к определенным районам населяют водоемы какого-нибудь одного типа. |о всех странах света обитают обыкновенная Щ^шя (Daphnia pulex), планктонная дафния
| « |
. longispina), босмина (Bosmina longirostris), kpyc (Chydorus sphaericus) и многие другие. Втоже время существуют виды, живущие только ■а Крайнем Севере (Daphnia arctica, Eurycercus ■acialis). Преимущественно в северной части СССР ■ Америки встречается своеобразный Holopedi-lim gibberum, имеющий поверх раковины студе-Иктую оболочку, придающую рачку вид прозрач-1юго шарика (табл. 48, 16). Этот рачок изредка Встречается и в средней части СССР, в водоемах, ■банных с моховыми болотами. Наконец, есть и ■ропические виды, в нашей стране обитающие на Витых водой рисовых полях Узбекистана, в За-■авказье, в водоемах дельты Волги. Высказыва-|ш предположение о том, что их покоящиеся ■яйца переносятся на колоссальные расстояния юздушными потоками.
I Некоторые ветвистоусые рачки населяют толь- т те водоемы, вода которых имеет кислую ре-■диюибедна минеральными солями, но большин-■jbo видов способно существовать в самых разнооб-, в том числе и неблагоприятных условиях,;ример в условиях недостаточного содержания юрода в воде. Немногочисленные морские представители вет-мусых ракообразных также распространены
очень широко, но приурочены преимущественно к прибрежным водам. Только в Каспийском море обитает несколько чрезвычайно свбеобразных видов (табл. 48, 6, 18), частично присущих только Каспию, частично встречающихся также в Азовском море и в опресненных районах Черного моря. Среди них особенно причудливы виды рода Сег-copagis, снабженные необыкновенно длинной хвостовой иглой, в 5—6 раз превосходящей по длине их тело. Каспийские ветвистоусые близки к некоторым пресноводным северным видам (например, Bythotrephes); предполагается, что при таянии великого ледника эти виды были внесены в Каспийское море и, приспособившись к новым для них условиям, дали начало каспийским эндемикам.
Значение ветвистоусых ракообразных в жизни стоячих пресноводных водоемов очень велико. Они служат излюбленным и весьма питательным кормом для многих рыб, как взрослых, так и молоди, и истребляются рыбами в огромном количестве. В кишечнике рипуса из уральского озера Увильда оказалось 47 тыс. босмин и 8,5 тыс. дафний. Чудской сиг, снеток, ряпушка, молодь судака, амурского сазана и многих других рыб питаются главным образом ветвистоусыми ракообразными. Питательность этого корма очень высока: содержание белка в теле дафнии достигает 50%, а жира 11 % по массе. Многие морские рыбы также в большом количестве поедают ветвистоусых.
Высокие пищевые качества ветвистоусых ракообразных в полной мере оценены рыбоводами. На рыбоводных заводах нашей страны осуществляется массовое разведение дафний и моин, которыми кормят молодь осетровых и лососевых рыб.
Перед рыбоводами, занимающимися этим трудным делом, стоит задача обеспечить беспрерывное партеногенетическое размножение разводимых рачков. Для этого необходимо создать в бассейнах, в которых разводятся рачки, постоянные благоприятные для них условия. В случае хотя бы временного ухудшения условий существования рачков их яйца, как было описано раньше, оплодотворяются и превращаются в покоящиеся. Подойдя к бассейну, рыбовод обнаруживает вместо массы живых копошащихся рачков плавающие на поверхности воды эфиппиумы, которых рыбки есть, конечно, не будут. Дело может кончиться гибелью от голода сотен и тысяч маленьких осет-рят и севрюжат.
Дафний и моин разводят в цементированных или просто выкопанных в земле бассейнах, воду которых удобряют навозом, отваром с жиротопни и азотобактерином. Эти вещества благоприятствуют массовому развитию бактерий, используемых в пищу рачками. Иногда бассейны делают проточными, что позволяет регулировать их температурный и газовый режим.
 ПОДКЛАСС РЕМИПЕДИИ (REMIPEDIA)
ПОДКЛАСС РЕМИПЕДИИ (REMIPEDIA)
В последние годы обследовано немало морских пещер. В них. обнаружено довольно много морских животных. Среди них особый интерес представляют рачки Speleonectes (рис. 199), настолько не похожие на других ракообразных, что они были выделены в самостоятельный подкласс. Тело этих животных, длиной до 43 мм, подразделено на голову, туловище, состоящее не более чем из 32 сегментов, и тельсон с фуркой. Голова, помимо обычных 2 пар антенн, несет парные лобные выросты. Ротовые конечности ноговидные, членистые, сходны по строению с первой парой туловищных ножек, вошедших в состав ротового аппарата. Туловищные ножки отходят от боков сегментов и состоят из 2 разделенных на членики ветвей. Животные плавают спиной вниз, загребая воду ножками. Эти слепые депигмеитированные животные сначала были обнаружены в затопленной морем пещере на глубине 19 м на Большом Багамском острове. Впоследствии в сходных местообитаниях их обнаружили на других островах Багамского архипелага. Самое удивительное, что они были найдены и в одной из лавовых пещер на Канарских островах. Архаическое однообразное расчленение этих рачков, строение их конечностей и разорванный тип ареала свидетельствуют о древности данной группы ракообразных. По-видимому, они представляют реликт фауны верхнемелового моря
Рнс. 199. Спелеонектес (Speleonectes).
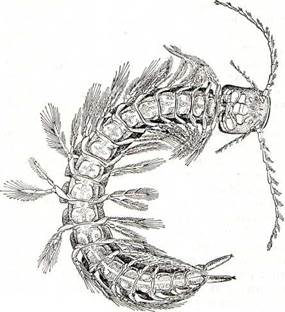
Тетис, один из заливов которого находился меш| Северо-Восточной Африкой и Северной Америке соединенными непрерывной береговой лини После расхождения материков и возникновеш Атлантического океана ареал этих животных «зался разорванным надвое. В настоящее врея известно 4 вида ремипедий, относимых к 3 рода!
ПОДЕЛАСС
ЦЕФАЛОКАРИДЫ
(CEPHALOCARIDA) I
В 1955 г. американский зоолог Сандер занимался изучением морской фауны близ Окш нографического института в Вудс-Холе, на бери Атлантического океана. Его интересовали не тол ко крупные, но и мелкие обитатели дна. На 1 большой глубине (9—20 м) он обнаружил малеи ких рачков, которые оживленно двигались среЛ частиц грунта, изгибая длинное тонкое тело. Вш мательно рассмотрев добычу, Сандерс установи: что эти рачки непохожи на всех известных до ci пор, но в то же время имеют те или иные общи признаки почти с каждым из известных подклж сов ракообразных. Это заставило изучить рачш более подробно, так как при исследовании i строения можно было надеяться выяснить родя венные отношения главных групп класса ракооо разных.
Эти рачки (длиной тела от 2,0 до 3,7 мм) имя крупную голову, задний край которой нависа! над первым грудным сегментом. Глаз у них нег Задние антенны расположены позади рта. У вес остальных ракообразных во взрослом состоят обе пары антенн находятся перед ртом, и толы у личинок они занимают такое же положени как у взрослых цефалокарид. За большой вер! ней губой открывается рот, по краям которог помещаются жвалы. Передние челюсти малея! кие, а задние имеют такое же строение, как Я дующие за ними грудные ножки: большая основ ная часть, снабженная внутренними выростам. служащая для передвижения, расчлененная вга ренняя ветвь и две листовидные наружные лопаси Сходство в строении и функциях челюстей и груи ных ножек может считаться признаком прими™ ной организации; у цефалокарид челюсти еще» достигли той степени специализации, которой ош достигли у других ракообразных.
Грудной отдел состоит из 8 сегментов. Грудш ножки совершают такие же движения, как у щ роногов. Они сгибаются последовательно, вызъш ток воды, направленный вперед, ко рту. При и гибании ножек назад вода с боков устремляете! между ними (рис. 200, 1 — 6), а при их изия
ш вперед вода проталкивается также вперед рпс 200, 7 — 9). Оседающие при этом на щетинки взвешенные частицы попадают в рот и поеда-ются. Конечности последнего грудного сегмента вдоразвиты или отсутствуют. | Брюшко уже и длиннее грудного отдела. В не-
■ входит 11 сегментов, оно заканчивается фур-
[шьными ветвями, вооруженными необычайно
шнными щетинками.
шокариды—гермафродиты. В яйцевом меш-к одновременно развивается только один (или
эмбрион, но зато некоторые виды, например incisa, размножаются круглый год. Из щ выклевываются личинки (метанауплиус), на-юминающие взрослых, но имеющие меньше сег-■нтов, их антенны и жвалы развиты полностью, ■обепары челюстей зачаточны (рис. 200, Г). Даль-кипее развитие происходит очень плавно: с каж-■1 последующей линькой добавляется 1—2 сег-инта тела, и так же постепенно увеличивается ■моконечностей. У одних видов через 13, у дру- т через 18 линек рачки достигают половозре-| мсти.
■Личинки, так же как и взрослые, ведут донный раз жизни. Они не похожи на науплиусов жаб-■ргих ракообразных, так как имеют сравнитель-
■ длинные расчлененные антенны, короткое ос-
иание задних антенн и двуветвистые жвалы. Эти
иризнаки свойственны науплиусам других рако-
■разных.
IB настоящее время описано 9 видов цефало-карид. Первый открытый Сандерсом вид цефало-Вшд — Hutchinsoniella macracantha — был собран на мелководье. Большинство последующих Ирдок также относятся к небольшой глубине. Ков 1962 г. Хесслер и Сандерс обнару-иш Н. macracantha у берегов Новой Англии на нубине 300 м, а в 1973 г. — новый вид Sander-plla bathyalis у побережья Юго-Западной Аф-[ на глубине даже 1227 —1559 м. |фалокариды встречены на восточном побе-рье Северной и Южной Америки, в заливе Сан-Франциско, в Карибском море, в Мексиканском шве. у побережий Западной Африки, Перу, рой Зеландии, у берегов Новой Каледонии и [южного побережья Японии. Можно предполо-ить, что впоследствии цефалокариды будут обнажены и в других районах Мирового океана. Сравнение признаков цефалокарид и других ркообразных показывает, что цефалокариды за-иают центральное положение в этом классе. I конечностей цефалокарид можно вывести все вш конечностей, свойственные разным подклас-|ши отрядам ракообразных. В особенности сход-вино форме, и по функциям конечности цефа-■карид и жаброногов. Строением головы и зад-■го конца тела цефалокариды напоминают вес-юношх, много общего у них и с личинками выс-ш ракообразных. По всей вероятности, цефа-
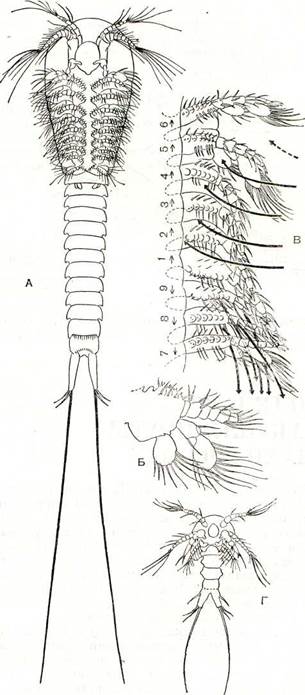
Рис. 200. Цефалокариды:
А — общий вид Hutchinsoniella; Б — ее грудная ношка; В — движение конечностей и вызываемые ими токи воды (обозначены стрелками, цифрами сверху показана последовательность взмахов ног); Г — первая личинка.
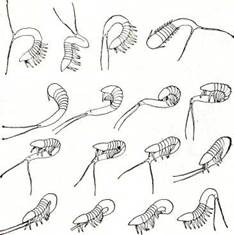
Рис. 201. Движения Hutchinsoniella на поверхности грунта.
локариды сохранили многие признаки, которыми обладали некогда существовавшие общие предки всех ракообразных.
ПОДКЛАСС
МАКСИЛЛОПОДЫ
(MAXILLOPODA)
Грудной отдел у Maxillopoda состоит из определенного числа сегментов, обычно из 6 или реже оно сокращено до 5 или 4. Грудные ножки служат только для передвижения или для возбуждения токов воды, но не несут дыхательной функции и лишены жевательных отростков. Ротовые придатки хорошо развиты, часто служат для фильтрации. Сложные глаза и жабры обычно отсутствуют, кровеносная система сильно упрощена или отсутствует. Развитие со сложным метаморфозом.
В состав этого подкласса наряду ео свободножи-вущими рачками, сохраняющими типичное для Maxillopoda строение, входят ракообразные, ведущие прикрепленный и паразитический образ жизни, сильно изменивший их организацию.
Maxillopoda распространены чрезвычайно широко. Они населяют как море, так и пресные воды, живут как на дне, так и в толще воды, обитают в подземных и в капиллярных водах, пропитывающих песок, опускаются на огромные океанические глубины, паразитируют на других животных, в том числе и на других ракообразных, — словом, они эволюционировали в различных направлениях и приспособились к существованию в самых разнообразных условиях.
В этот подкласс включают 5 отрядов — весло ногих, карпоедов, мистакокарид, мешкогрудых i усоногих.
ОТРЯД ВЕСЛОНОГИЕ РАКООБРАЗНЫЕ (COPEPODA)
При изучении жизни каждого водоема — от 1 ленькой лужи до океана — исследователь ветре чается с этими рачками. Планктонная сеть, протя нутая на любой глубине океана, в жарких тр«пиках или в холодных приполярных водах, прино сит улов, состоящий преимущественно из вес.» ногих рачков. В ручьях и озерах глубоких темнш пещер также обитают эти ракообразные. Они Mat; сами плавают во всех обычных пресноводных ш доемах, часто не уступая по численности ветви тоусым. Некоторые из них паразитируют на рн бах и других водных животных, причем нередк под влиянием паразитизма меняются настолько. что не сразу можно угадать их истинную природ; Веслоногие массами поедаются рыбами. Сельдь иваси и многие другие рыбы питаются почти | ключительно веслоногими. Большинство этих pal кообразных — животные микроскопические, длина их тела от 1 мм и меньше до 5 мм; лишь отдел ные глубоководные и паразитические виды доел гают длины 1—3 см. В настоящее время насчии-вается около 7500 видов веслоногих.
Тело свободноживущих веслоногих разделяете! на голову, грудь и брюшко (рис. 202). Голова слит! ная, без всяких следов сегментации, срастаеш с первым грудным сегментом. Передний коня головы часто вытянут в загнутый вниз клюв, щ рострум. Очень характерно отсутствие парни фасеточных глаз; на лобной части головы распм ложен только науплиальный глазок. Имени это обстоятельство позволило датскому натурам ту Мюллеру в свое время назвать обычш пресноводных веслоногих «циклопами», по имея одноглазых гигантов из греческой мифологии. I
Голова снабжена 5 парами придатков. Передни антенны часто очень длинные, иногда длин! тела, и участвуют в плавании и парении рачк» Кроме того, они выполняют и функции органа! чувств: на них сидят чувствительные щетинкиi цилиндрические чувствительные придатки. Задние антенны короткие, обычно двуветвистые. Жва-лы мощные и имеют двуветвистый щупик. Их аи вательная сильно хитинизированная часть 1 ладает острыми зубцами, помогающими раздроблять пищу. При внимательном рассмотрении зуб-цов жвал некоторых морских веслоногих выжни л ось, что эти зубцы покрыты кремневыми и ронками, увеличивающими их прочность (рис. 203) Открытие кремневых коронок интересно в Щ отношениях. Во-первых, оно указывает на способ ность веслоногих ракообразных усваивать и щ
центрировать кремний; такой способности лишены почти все высшие беспозвоночные — черви, мол-лики, а также другие членистоногие. Во-вторых, южно надеяться найти в геологических отложениях кремневые коронки древних веслоногих, почти совсем не сохранившихся в ископаемом состоянии.
Передние челюсти веслоногих устроены очень можно, так как снабжены внутренними и наружными лопастями и многочисленными перистыми щетинками. Задние челюсти имеют только внутренние лопасти и также многочисленные щетинки. К головным придаткам присоединяется пара оцноветвистых ногочелюстей, принадлежащих переднему слившемуся с головой грудному сег-шнту.
Задние антенны, щупики жвал и передние че-Iлюети фильтрующих веслоногих совершают час-ше и беспрерывные взмахи, создающие круговороты воды, которые приносят взвешенные пищевые частицы. Эти частицы отфильтровываются главным образом щетинками задних челюстей. Грудной отдел состоит из 5 сегментов с ясно ви-рмыми границами между ними. Все 5 пар грудных ножек у примитивных веслоногих устроены одинаково. Каждая ножка состоит из 2-членико-вой основной части и двух обычно 3-члениковых ветвей, вооруженных шипами и щетинками. Эти ножки совершают одновременные взмахи, действуя как весла и отталкивая тело рачка от воды. У многих более специализированных видов пятая пара ножек самца преобразована в аппарат, приспособленный для удерживания самки во вре-ш спаривания и прикрепления сперматофоров [нееполовым отверстиям. Нередко пятая пара ног редуцирована.
Брюшной отдел состоит из 4 сегментов, но у iсамок часто их число меньше, так как некоторые из них сливаются между собой. На переднем брюшном сегменте открывается парное или не-варное половое отверстие, и у самки этот сегмент кто крупнее остальных. Брюшко заканчивается мьсоном, с которым сочленены фуркальные [ветви. Каждая из них вооружена несколькими ■очень длинными, иногда перистыми щетинками. Эти щетинки особенно сильно развиты у планк-юнных видов, у которых они увеличивают поверхность тела (по отношению к его объему), чем способствуют парению рачка в толще воды. | Строение паразитических видов сильно откло-иется от описанного. Их головные придатки пре-рцаются в органы прикрепления к хозяину и iсвязи с этим часто имеют вид хитиновых крючков, погруженных в кожу рыб или других водных животных. Плавательные ножки иногда исчезают совсем, или число их сокращается. В большей или мыней степени исчезает сегментация, покровы становятся мягкими. Специализированные парада приобретают червеобразную форму тела и
теряют всякое сходство с ракообразными. Однако можно подобрать полную серию переходов между такими предельно упрощенными видами и нормальными свободноживущими. Кроме того, яйца прикрепляются к телу паразитических видов совершенно так же, как к телу свободноживущих, и из яиц выходят такие же личинки.
Дыхание веслоногих осуществляется всей поверхностью тела, жабры отсутствуют. С этим, возможно, связано и слабое развитие или даже отсутствие кровеносной системы. Сердце есть только у представителей подотряда Calanoida, да и у них оно невелико, хотя и бьется очень часто: например, у морского рачка Labidocera оно совершает более 150 ударов в 1 мин. У остальных веслоногих полостная жидкость приводится в движение сокращениями кишечника.
| Рис. 202. Общий вид самок трех подотрядов веслоногих: 1 — Calanoida; 2 — Cyclopoida; 3 — Harpacticoida. |
| Рис. 203. Край жующей пластинки Eucalanus bungii и один зубец при большом увеличении. |
Все веслоногие раздельнополы. Самцы обычно меньше самок и у свободноживущих видов отличаются от самок искривленными (одной или обеими) передними антеннами, а также иным строением пятой пары грудных ножек. Особенно резко различны самцы и самки многих паразитических видов. У некоторых из них прикрепленные к хозяи-
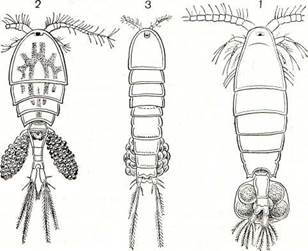


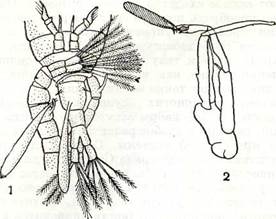
Рис. 204. Спаривание Calanoida:
1 — прикрепление сперматофора к генитальному сегменту самки у Diaptoraus; 2 — пятая пара ног Pareuchaeta gracialis, последний членик левой ноги со «щипчиками», удерживающими сперматофор.
ну самки полностью теряют сегментацию и конечности, а маленькие самцы сохраняют облик поздней личинки и свободно плавают. У других (семейство Lernaeopodidae) маленькие самцы, также сходные с поздней личинкой, своими клешневидными челюстями и ногочелюстями прикрепляются к крупным червеобразным паразитическим самкам.
При спаривании самец удерживает самку пятой парой грудных ножек и первыми антеннами и при помощи той же пятой пары ног приклеивает колбасовидный сперматофор близ ее половых отверстий, т. е. к нижей стороне первого брюшного сегмента. У некоторых видов одна из ветвей пятой пары ног самца снабжена на конце щипчиками, захватывающими сперматофор и переносящими его на нужное место (рис. 204). Из сперматофора сперма попадает в семяприемник самки. При выметывании яиц они оплодотворяются.
Большинство морских планктонных видов откладывает яйца прямо в воду, но у пресноводных и паразитических, а также у морских донных и прибрежных видов яйца склеиваются между собой специальным секретом и прикрепляются близ половых отверстий самки. У одних веслоногих таким образом формируется один, у других — два яйцевых мешка, которые самка носит на себе до выхода из яиц личинок.
Из яйца выходит личинка науплиус. Личинка многократно линяет и постепенно приближается по своим признакам к взрослому рачку. Различают 11—12 личиночных стадий: 5—6 науплиаль-ных и 6 копеподитных, из которых последняя — взрослая половозрелая стадия. Сначала личинки имеют лишь две пары антенн и жвалы, затем у них развиваются остальные ротовые придатки, но тело остается несегментированным. Копеподитпые стадии отличаются сегментацией заднего конца
тела и постепенным развитием грудных ноли Для завершения метаморфоза разным веслоног требуется различное время, да и биология личив далеко не у всех видов одинакова.
Образ жизни, способ питания и среда обитая веслоногих рачков настолько разнообразны, i лучше рассмотреть этот отряд не целиком, а к»; дый из входящих в него подотрядов в отдельной В пределах отряда веслоногих принято раза чать 13 подотрядов, из которых 10 объедини исключительно паразитические виды. Свой ноживущие веслоногие относятся к подотряд Calanoida, Cyclopoida и Harpacticoida (рис. 20! Calanoida — исключительно планктонные я вотные. Их голова и грудь значительно длиш узкого брюшка, передние антенны очень дли ные, превосходят голову и грудь, а нередко и'У тело рачка, если есть яйцевой мешок, то один. Cyclopoida обитают преимущественно на I или в придонных слоях воды. Среди них есн паразиты. Их голова и грудь относительно все тела гораздо короче, чем у Calanoida, переда антенны короткие, не достигают и середины га вогрудного отдела, брюшко уже груди. Саш всегда носят два яйцевых мешка.
Harpacticoida, за единичными исключения! живут на дне и больше ползают, чем плава! Тело их червеобразно благодаря тому, что бри ной отдел почти не отличается по ширине от rpj ного. Передние антенны очень короткие, cm большинства видов образуют один яйцевой ] шок.
Представители всех трех подотрядов населя как моря, так и пресные воды.
ПОДОТРЯД КАЛАНИДЫ (CALANOIDA)
Вся организация Calanoida превосходно щ способлена к жизни в толще воды. Длинные I тенны и перистые щетинки фуркальных вей позволяют морскому Calanus или пресноводнок Diaptomus неподвижно парить в воде, лишь очи медленно погружаясь. Этому способствуют наход! щиеся в полости тела рачков капли жира, умев шающие их плотность. Во время парения щ рачка располагается вертикально или наклони причем передний конец тела расположен вш заднего. Опустившись на несколько сантиметр вниз, рачок делает резкий взмах всеми груднш ножками и брюшком и возвращается на прежш уровень, после чего все повторяется сначала, Tj ким образом, путь рачка в воде рисуется зигзак образной линией (рис. 205, 1). Некоторые морей Calanoida, как, например, приповерхностный bi ярко-синего цвета Pontellina mediterranea, j вершают настолько резкие скачки, что выпри? гивают из воды и пролетают какое-то простри ство по воздуху наподобие летучих рыб.








