 2015-08-21
2015-08-21 3707
3707Для суккулентов (роды Crassula, Bryophyilum и др.) характерен суточный цикл метаболизма С4-кислот с образованием яблочной кислоты ночью. От английского выражения crassulacean acid metabolism этот тип фотосинтеза сокращенно называют СAM-метаболизм. Устьица этих растений днем обычно закрыты, что предотвращает потерю воды, и открываются ночью. СО2 поступает в листья, где при участии содержащейся в цитоплазме ФЕП-карбоксилазы взаимодействует с фосфоенолпируватом, образуя оксалоацетат. Источником ФЕП служит крахмал. То же самое происходит и с СО2, который освобождается в клетках в процессе дыхания. Образовавшийся оксалоацетат восстанавливается под действием NADH-зависимой малатдегидрогеназы до яблочной кислоты, которая накапливается в вакуолях клеток листа. Это приводит к закислению клеточного сока в ночное время.
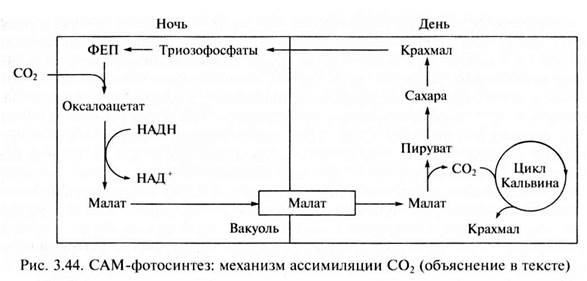
Днем в условиях высокой температуры, когда устьица закрыты, малат транспортируется из вакуолей в цитоплазму и там декарбоксилируется при участии малатдегидрогеназы декарбоксилирующей (малик-энзима) с образованием СО2 и пирувата. СО2 поступает в хлоропласты и включается в них в цикл Кальвина, участвуя в синтезе сахаров.
Таким образом, у растений с фотосинтезом по типу толстянковых много общего с С4-путем фотосинтеза. Однако при САМ-метаболизме фиксация СО2 с образованием малата (ночью) и декарбоксилирование малата с высвобождением СО2 и пирувата (днем) разделены во времени. При достаточном количестве воды ряд растений с метаболизмом по типу толстянковых могут вести себя как С3-растения. В свою очередь некоторые растения с С3-путем фотосинтеза при недостатке воды проявляют черты САМ-метаболизма.
5. Фотодыхание ( гликолатный путь, или С2-путь)
Фотодыхание − активируемый светом процесс высвобождения СО2 и поглощения О2 (отличается от «темнового» дыхания митохондрий). Так как при этом первичным продуктом является гликолевая кислота, то этот путь получил название гликолатного. У С3-растений с малой эффективностью фотосинтеза интенсивность фотодыхания может достигать 50% от интенсивности фотосинтеза.
Фотодыхание
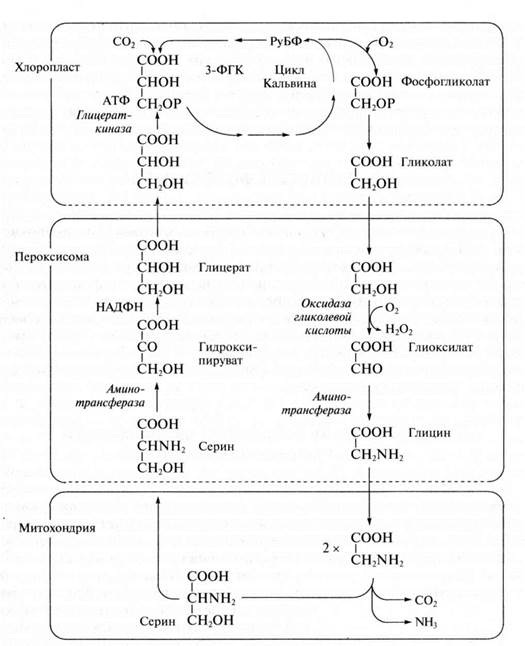
Фотодыхание у С3-растений усиливается при низком содержании СО2 и высоких концентрациях О2. В этих условиях РДФ-карбоксилаза (рибулозодифосфат карбоксилаза) в хлоропластах может функционировать как оксигеназа, катализируя окислительное расщепление рибулозо-1,5-дифосфата на 3-ФГК и 2-фосфогликолевую кислоту, которая затем дефосфорилируется в гликолевую кислоту. Молекулы СО2 и О2 конкурируют между собой в каталитическом центре РДФ-карбоксилазы: при относительно высоких концентрациях СО2 и низких О2 преобладает карбоксилирование, а высокие концентрации О2 и низкое содержание СО2 благоприятствуют окислению и образованию фосфогликолевой кислоты. В том же направлении действует и повышение температуры.
Фотодыхание осуществляется в результате взаимодействия хлоропластов, пероксисом и митохондрий. Гликолат из хлоропласта поступает в пероксисому и там окисляется гликолатоксидазой до глиоксилата. Возникающая перекись водорода устраняется каталазой пероксисомы. Глиоксилат аминируется, превращаясь в глицин (качестве донора аминогруппы функционирует глутамат).
Глицин транспортируется в митохондрию. Здесь из двух молекул глицина образуется серии и освобождается СО2. Теперь цикл замыкается: серии может снова поступать в пероксисому и там передать свою аминогруппу на пируват. При этом из пирувата возникает аланин, из серина — гидрокси-пируват, который сразу после этого восстанавливается в глицерат. Затем глицерат может снова попасть в хлоро-пласты и благодаря фосфорилированию включиться в цикл Кальвина.
Последовательность реакций фотодыхания не обязательно образует цикл. Гликолатный путь С3-растений может завершиться в митохондриях. Конечными продуктами в этом случае являются серин и СО2. Освобождение СО2 объясняет, почему нетто-фотосинтез (чистая продуктивность) при интенсивном фотодыхании снижается.
У С4-растений СО2, выделяющийся в результате фотодыхания, перехватывается в клетках мезофилла, где из ФЕП и СО2 образуются оксалоацетат и малат. Затем малат «отдает» свой СО2 хлоропластам обкладки, где функционирует цикл Кальвина. В связи с этим становится понятным высокий нетто-фотосинтез С4-растений.
Физиологическое значение фотодыхания: гликолатный путь приводит к синтезу глицина и серина, в пероксисомах происходит восстановление NADP+; в процессе образования серина может генерироваться АТР. Показано, что С3-растения, помещенные в атмосферу с низким парциальным давлением О2 и высокой концентрацией СО2, ведут себя подобно С4-растениям, т. е. имеют низкий уровень фотодыхания.
Вывод: термин «фотодыхание» имеет лишь формальный смысл (О2 потребляется, СО2 выделяется), но к дыханию этот процесс прямого отношения не имеет.








