 2020-06-12
2020-06-12 994
994В отложениях этого периода находят огромные залежи каменного угля. Отсюда и произошло название периода. Есть и другое его наименование – карбон.
Каменноугольный период подразделяется на три отдела: нижний, средний и верхний. В этот период физико‑географические условия Земли претерпели значительные изменения. Неоднократно менялись очертания материков и морей, возникали новые горные хребты, моря, острова. В начале карбона происходит значительное опускание суши. Морем были затоплены обширные площади Атлантии, Азии, Гондваны. Уменьшилась площадь больших островов. Исчезли под водой пустыни северного континента. Климат стал очень теплым и влажным.
В нижнем карбоне начинается интенсивный горообразовательный процесс: формируются Арденны, Гарц, Рудные горы, Судеты, Атласские горы, Австралийские Кордильеры, Западно‑Сибирские горы. Море отступает.
В среднем карбоне суша вновь опускается, но гораздо меньше, чем в нижнем. В межгорных котловинах накопляются мощные толщи континентальных отложений. Формируется Восточный Урал, Пеннинские горы.
В верхнем карбоне море вновь отступает. Значительно уменьшаются внутриконтинентальные моря. На территории Гондваны возникают большие ледники, в Африке и Австралии – несколько меньшие.
В конце карбона в Европе и Северной Америке климат претерпевает изменения, становится частично умеренным, а частично жарким и сухим. В это время происходит формирование Центрального Урала.
Морские осадочные отложения каменноугольного периода представлены главным образом глинами, песчаниками, известняками, сланцами и вулканогенными породами. Континентальные – преимущественно углем, глинами, песками и другими породами.
Усиленная вулканическая деятельность в карбоне привела к насыщению атмосферы углекислым газом. Вулканический пепел, являющийся чудесным удобрением, сделал плодородными карбоновые почвы.
Теплый и влажный климат господствовал на материках длительное время. Все это создало чрезвычайно благоприятные условия для развития наземной флоры, в том числе высших растений каменноугольного периода – кустов, деревьев и травянистых растений, жизнь которых была тесно связана с водой. Они росли главным образом среди громадных болот и озер, близ солоноватоводных лагун, на побережье морей, на влажной илистой почве. По образу жизни они походили на современных мангровых, которые растут на низинных берегах тропических морей, в устьях больших рек, в болотистых лагунах, поднимаясь над водой на высоких корнях‑ходулях.
Значительное развитие в каменноугольный период получили плауновидные, членистостебельные и папоротниковые, давшие большое количество древовидных форм.
Древовидные плаунообразные достигали 2 м в диаметре и 40 м в высоту. У них еще не было годичных колец. Пустой ствол с мощной разветвленной кроной надежно удерживался в рыхлой почве большим корневищем, разветлявшимся на четыре главные ветви. Эти ветви в свою очередь дихотомически разделялись на корневые отростки. Их листья, до метра в длину, густыми султанообразными пучками украшали концы ветвей. На концах листьев располагались почки, в которых развивались споры. Стволы плаунообразных были покрыты чешуей – рубцами. К ним прикреплялись листья. В этот период были распространены гигантские плаунообразные – лепидодендроны с ромбическими рубцами на стволах и сигиллярии с шестигранными рубцами. В отличие от большинства плаунообразных у сигиллярий был почти неразветвленный ствол, на котором вырастали спорангии. Среди плаунообразных были и травянистые растения, полностью вымершие в пермский период.
Членистостебельные растения разделяются на две группы: клинолистые и каламиты. Клинолистые являлись водными растениями. У них был длинный, членистый, слегка ребристый стебель, к узлам которого кольцами прикреплялись листья. Почковидные образования содержали споры. На воде клинолистые держались с помощью длинных разветвленных стеблей, похожих на современный водяной лютик. Клинолистые появились в среднем девоне и вымерли в пермский период.
Каламиты были древовидными растениями до 30 м высотой. Они образовывали болотные леса. Некоторые виды каламитов проникли далеко на материк. Их древние формы имели дихотомические листья. Впоследствии преобладали формы с простыми листьями и годичными кольцами. Эти растения имели сильно разветвленное корневище. Нередко из ствола вырастали дополнительные корни и ветви, покрытые листьями.
В конце карбона появляются первые представители хвощевых – небольшие травянистые растения. Среди карбоновой флоры видную роль играли папоротники, в частности травянистые, по своему строению напоминавшие псилофитов, и настоящие папоротники – большие древовидные растения, корневищем закреплявшиеся в мягком грунте. У них был шершавый ствол с многочисленными ветвями, на которых росли широкие папоротниковидные листья.
Голосеменные карбоновых лесов относятся к подклассам семенных папоротников и стахиоспермид. Их плоды развивались на листьях, что является признаком примитивной организации. В то же время у линейных или ланцетных листьев голосеменных было довольно сложное жилкование. Наиболее совершенные растения карбона – кордаиты. Их цилиндрические безлистые стволы до 40 м высотой разветвлялись. Ветви имели на концах широкие линейные или ланцетные листья с сетчатым жилкованием, Мужские спорангии (микроспорангии) имели вид почек. Из женских спорангиев развивались ореховидные плоды. Результаты микроскопического исследования плодов показывают, что эти растения, похожие на цикадовых, являлись переходными формами к хвойным растениям.
В каменноугольных лесах появляются первые грибы, моховидные растения (наземные и пресноводные), образовывавшие иногда колонии, и лишайники.
В морских и пресноводных бассейнах продолжают существовать водоросли: зеленые, красные и харовые.
При рассмотрении каменноугольной флоры в целом поражает разнообразие форм листьев древовидных растений. Рубцы на стволах растений на протяжении всей жизни держали длинные, ланцетные листья. Концы ветвей были украшены огромными лиственными кронами. Иногда листья росли по всей длине ветвей.
Другой характерный признак каменноугольной флоры – развитие подземной корневой системы. Сильно разветвленные корни разрастались в илистой почве и из них произрастали новые побеги. Порой значительные площади были изрезаны подземными корнями.
В местах быстрого накопления илистых осадков корни удерживали стволы многочисленными побегами. Важнейшая особенность каменноугольной флоры заключается в том, что растения не отличались ритмическим ростом в толщину.
Распространение одних и тех же каменноугольных растений от Северной Америки до Шпицбергена свидетельствует о том, что от тропиков до полюсов господствовал относительно равномерный теплый климат, на смену которому в верхнем карбоне пришел довольно прохладный. В прохладном климате росли голосеменные папоротники и кордаиты.
Рост каменноугольных растений почти не зависел от времен года. Он напоминал рост пресноводных водорослей. Времена года, вероятно, мало отличались друг от друга.
При изучении каменноугольной флоры можно проследить эволюцию растений. Схематически она выглядит следующим образом: бурые водоросли – папоротники – псилофиты – птеридоспермиды (семенные папоротники) – хвойные.
Отмирая, растения каменноугольного периода падали в воду, их заносило илом, и, пролежав миллионы лет, они постепенно превратились в уголь. Каменный уголь образовывался изо всех частей растения: древесины, коры, ветвей, листьев, плодов. В уголь превращались и остатки животных. Об этом свидетельствует тот факт, что остатки пресноводных и наземных животных в карбоновых отложениях встречаются относительно редко.
Морской животный мир карбона характеризовался разнообразием видов. Чрезвычайно распространенными были фораминиферы, в частности фузулиниды с веретеновидными раковинами величиной с зерно.
В среднем карбоне появляются швагерины. Их шаровидная раковина была величиной с небольшую горошину. Из раковин фораминифер позднего карбона в отдельных местах образовались залежи известняков.
Среди кораллов еще встречались немногочисленные роды табулят, но начинали преобладать хететиды. Одиночные кораллы нередко имели толстые известковые стенки. Колониальные кораллы образовывали рифы.
В это время интенсивно развиваются иглокожие, в частности морские лилии и морские ежи. Многочисленные колонии мшанок иногда образовывали мощные известняковые отложения.
Чрезвычайно развились плеченогие моллюски, в частности продуктусы, по приспособляемости и географическому распространению намного превосходившие всех встречающихся на Земле плеченогих. Величина их раковин достигала 30 см в поперечнике. Одна створка раковины была выпуклой, а другая – в виде плоской крышечки. Прямой удлиненный замочный край часто имел полые шипы. У некоторых форм продуктусов шипы в четыре раза превышали диаметр раковины. С помощью шипов продуктусы держались на листьях водных растений, переносивших их по течению. Иногда своими шипами они прикреплялись к морским лилиям или водорослям и жили возле них в висячем положении. У рихтофений одна створка раковины преобразовалась в рог длиной до 8 см.
В каменноугольный период почти полностью вымирают наутилоидеи, за исключением наутилусов. Данный род, распавшийся на 5 групп (которые были представлены 84 видами), дожил до нашего времени. Продолжают существовать ортоцерасы, раковины которых имели резко выраженное внешнее строение. Роговидно согнутые раковины циртоцерасов почти не отличались от раковин их девонских предков. Аммониты были представлены двумя отрядами – гониатитами и агониатитами, как и в девонском периоде; двустворчатые моллюски – одномышечными формами. Среди них – множество пресноводных форм, населявших карбоновые озера и болота.
Появляются первые наземные гастроподы – животные, дышавшие легкими.
Трилобиты достигли значительного расцвета в ордовикский и силурийский периоды. В каменноугольный период сохранились лишь немногочисленные их роды и виды.
К концу каменноугольного периода трилобиты почти полностью вымерли. Этому способствовало то, что головоногие моллюски и рыбы питались трилобитами и потребляли ту же пищу, что и трилобиты. Строение тела трилобитов было несовершенным: панцирь не защищал брюха, конечности были небольшими и слабыми. Трилобиты не имели органов нападения. Некоторое время они могли защищаться от хищников, свертываясь наподобие современных ежей. Но в конце карбона появились рыбы с мощными челюстями, разгрызавшими их панцирь. Поэтому от многочисленного типа в перми сохранился только один род.
В озерах каменноугольного периода появляются ракообразные, скорпионы, насекомые.
Карбоновые насекомые обладали признаками многих родов современных насекомых, поэтому отнести их к какому‑либо одному известному нам теперь роду невозможно. Несомненно, предками насекомых каменноугольного периода были ордовикские трилобиты. У девонских и силурийских насекомых было много общего с некоторыми их предками. Они играли уже значительную роль в животном мире.
Однако подлинного расцвета насекомые достигли в каменноугольном периоде. Представители самых малых из известных видов насекомых были 3 см в длину; размах крыльев у самых крупных (например, у стенодиктии) достигал 70 см, у древней стрекозы меганевры – одного метра. Тело меганевры имело 21 сегмент. Из них 6 составляли голову, 3 – грудь с четырьмя крыльями, 11 – брюшко, конечный сегмент походил на шиловидное продолжение хвостового щита трилобитов. Многочисленные пары конечностей были расчленены. С их помощью животное и ходило, и плавало. Молодые меганевры обитали в воде, превращаясь во взрослых насекомых в результате линьки. У меганевры были крепкие челюсти и сложные глаза.
В верхнем каменноугольном периоде древние насекомые вымерли, их потомки были более приспособленными к новым условиям жизни. Прямокрылые в ходе эволюции дали термитов и стрекоз, эвриптерусы – муравьев. Большинство древних форм насекомых переходили к наземному образу жизни лишь в зрелом возрасте. Размножались они исключительно в воде. Таким образом, перемена влажного климата на более сухой для многих древних насекомых была катастрофой.
В каменноугольном периоде появляется много акул. Это были еще не настоящие акулы, населяющие современные океаны, однако по сравнению с другими группами рыб они представляли собой самых совершенных хищников. В отдельных случаях их зубы и плавниковые шипы переполняют каменноугольные отложения. Это свидетельствует о том, что каменноугольные акулы жили в любой воде. Зубы зазубренные, широкие, режущие, бугристые, так как акулы питались разнообразнейшими животными. Постепенно они истребили примитивных девонских рыб. Ножевидные зубы акул легко разгрызали панцирь трилобитов, широкие бугристые зубные пластинки хорошо раздробляли толстые раковины моллюсков. Пиловидные, заостренные ряды зубов позволяли акулам питаться колониальными животными. Формы и размеры акул были такими же разнообразными, как и способ их питания. Одни из них окружали коралловые рифы и с молниеносной быстротой преследовали свою добычу, другие же неторопливо охотились на моллюсков, трилобитов либо зарывались в ил и подстерегали добычу. Акулы с пиловидным выростом на голове искали жертв в зарослях морских водорослей. Крупные акулы часто нападали на более мелких, поэтому у некоторых из последних для защиты в ходе эволюции возникли плавниковые шипы и накожные зубы.

Акула из рода плевракант.
Акулы интенсивно размножались. Это в конечном итоге привело к перенаселению моря этими животными. Были истреблены многие формы аммонитов, исчезли одиночные кораллы, представлявшие для акул легкодоступную питательную пищу, значительно сократилось число трилобитов, погибли все моллюски, имевшие тонкую раковину. Лишь толстые раковины спириферов не поддавались хищникам. Сохранились и продуктусы. Они защищались от хищников длинными шипами.
В пресноводных бассейнах каменноугольного периода жило множество эмалевочешуйчатых рыб. Некоторые из них прыгали по илистому берегу, наподобие современных рыб‑прыгунов. Спасаясь от врагов, насекомые покинули водную среду и заселили сушу сначала вблизи болот и озер, а затем – горы, долины и пустыни каменноугольных материков.
Среди насекомых каменноугольного периода отсутствуют пчелы и бабочки. Это понятно, так как в то время еще не было цветковых растений, чьей пыльцой и нектаром питаются эти насекомые.

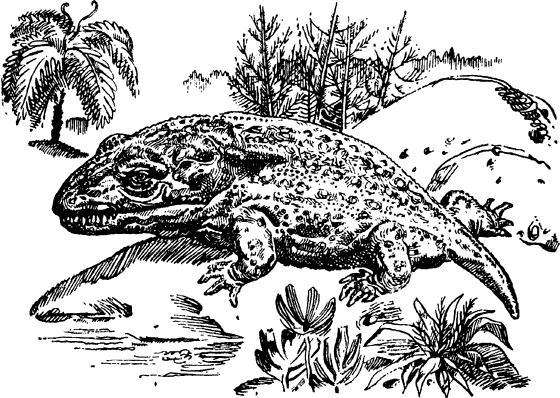
Урокодилусы – земноводные каменноугольного периода.
Животные, дышащие легкими, впервые появляются на материках девонского периода. Ими были земноводные. Жизнь земноводных тесно связана с водой, поскольку размножаются они лишь в воде. Теплый влажный климат карбона чрезвычайно благоприятствовал расцвету земноводных. Их скелеты еще не полностью окостенели, челюсти имели нежные зубчики. Кожа была покрыта чешуей. За низкий крышеобразный череп вся группа земноводных получила наименование стегоцефалов (панцироголовых). Размеры тела земноводных колебались от 10 см до 5 м. У большинства из них было четыре ноги с короткими пальцами. Некоторые имели когти, позволявшие им лазить по деревьям. Появляются и безногие формы. В зависимости от образа жизни земноводные приобрели тритоновидную, змеевидную, саламандровидную формы. В черепе земноводных было пять отверстий: два носовых, два глазных и теменной глаз. Впоследствии этот теменной глаз преобразовался в шишковидную железу головного мозга млекопитающих. Спина у стегоцефалов была голой, а брюхо покрывала нежная чешуя. Они населяли неглубокие озера и болотистые места близ побережья.

Эдафозавр.
Наиболее характерный представитель первых пресмыкающихся – эдафозавр. Он напоминал огромную ящерицу. На спине у него имелся высокий гребень из длинных костяных шипов, соединенных между собой кожистой перепонкой. Эдафозавр был травоядным ящером и жил вблизи каменноугольных болот.
С каменноугольными отложениями связано большое количество каменноугольных бассейнов, залежи нефти, железа, марганца, меди, известняков.
Длился этот период 65 млн. лет.
Пермский период
Пермские отложения впервые были описаны вблизи города Перми (РСФСР). Пермский период подразделяют на два отдела: нижний и верхний.
В результате горообразовательных процессов возникла Урало‑Тянь‑Шаньская платформа, Атлантия соединилась с Азией. Образовался огромный северный материк Лавразия. Продолжают формироваться Американские Кордильеры и Анды, Австралийские Кордильеры и Донецкий кряж. На территории Гондваны образовались большие углубления, в которых накапливались толщи осадочных пород: песков, гипсов, конгломератов.
Климат стал очень сухим. Исчезают многие внутриконтинентальные моря, озера, болота. Высокие горы обусловили дифференциацию климатических условий. В результате деятельности многочисленных вулканов возникли толщи вулканических пород.
Флора первой половины пермского периода отличается от флоры каменноугольного тем, что в это время значительно уменьшилось количество сигиллярий, лепидодендронов и кордаитов, преобладающими же формами стали папоротники и новые группы голосеменных растений. В болотах и мелких заливах, как и в каменноугольном периоде, еще растут каламиты, а поблизости – древовидные и травянистые папоротники.
Распространяются хвойные, гинкговые и саговники. По своему внешнему виду саговники напоминают пальмы. Органы размножения саговников – мужские и женские шишки. Их семена небольшие.
Из гинкговых до наших дней дожил лишь один вид. Это большое дерево с лопастевидными листьями. Сохранились гинкго благодаря человеку. Китайцы и японцы считали это дерево священным и выращивали вблизи храмов. Теперь гинкго растет почти во всех ботанических садах.
Особого развития в пермский период достигли языковые папоротники. На пучке корней, закрепленном в мягкой почве многочисленными отростками, поднимался шершавый ствол с ветвями, на которых располагались широкие папоротникообразные листья. Среди языковых папоротников различают кустовые и древовидные формы. На поперечных сечениях некоторых стволов окаменелых деревьев обнаружены кольца, свидетельствующие о сезонных изменениях климата. Неравномерность развития колец древесины могла быть связана как с более холодным временем года, так и с засушливым сезоном на протяжении одного или даже нескольких лет.
По своему виду хвойные растения напоминали современные араукарии, растущие на островах вблизи Австралии. Росли также кордаиты, близкие к современным. Их далекие потомки произрастают в Новой Зеландии. Кордаиты были очень похожи на сосну, остатки которой также встречаются в пермских отложениях. Из этих растений на протяжении пермского периода на побережьях, в долинах рек, в болотах и других влажных местах формировались мощные толщи каменного угля.
Растения и животные с известковыми раковинами поглотили из атмосферы каменноугольного периода огромное количество углекислого газа. Одновременно растения обогатили атмосферу свободным кислородом. В пермском периоде состав атмосферы приближался к современному, возникли климатические зоны. Расцвет травянистой растительности создал условия для возникновения и развития травоядных форм животных.
В прибрежных районах Гондванского материка, который простирался от Бразилии поперек Атлантического океана, включал Америку и через Мадагаскар достигал Индии и Австралии, находятся мощные толщи красных и желтых песчаников с остатками морской фауны: губок, двустворчатых, головоногих, брюхоногих, плеченогих моллюсков, иглокожих. Однако морской животный мир пермского периода намного беднее каменноугольного.
Фораминиферы встречаются редко, резко уменьшается количество губок, кораллов, иглокожих. Почти отсутствуют трилобиты. В конце перми вымирают табуляты, древние ежи.
Среди головоногих моллюсков развились новые формы аммонитов. В общем головоногие моллюски в пермских морях приобретают очень сложную организацию. Появляются наутилоидеи. Возникают формы брахиопод, живущие в наше время в Индийском океане. В пресных и солоноватых водах встречаются двустворчатые моллюски. Среди брюхоногих моллюсков развились главным образом переднежаберные. Продолжают существовать мшанки. Они образовывали рифы. Значительного развития достигли рачки остракоды и червеобразные.
Среди позвоночных в пермских морях значительное место занимали акулообразные – хрящевые рыбы со скрученными в спираль зубами. Появляются пресноводные акулы. Уменьшается количество кистеперых рыб. С началом пермского периода земноводные становятся довольно разнообразными. Небольшие формы, величиной в несколько сантиметров, жили рядом с гигантскими предками лягушек, достигавшими размеров быка.
Земноводные различались не только по размерам, но и по образу жизни. Обитали они и в воде, и на суше, питались насекомыми и рыбами, водорослями и папоротниками. Приспосабливаясь к наземным условиям, они все меньше времени проводили в воде.
Пресмыкающиеся по форме и строению тела сильно напоминали стегоцефалов. Однако самые примитивные представители данного класса были уже гораздо лучше приспособлены к условиям жизни на суше (имеется в виду способ размножения и развития зародышей). В то время как земноводные, подобно своим предкам – рыбам, размножались, откладывая икру в воду, пресмыкающиеся стали класть яйца непосредственно на суше. Более крупные, чем икра, яйца обладали значительным запасом питательных веществ, что позволяло зародышу развиваться минуя стадию личинки. Детеныши пресмыкающихся отличались от взрослых лишь по величине, личинки же земноводных вели водный образ жизни, отличаясь по своему строению от взрослых особей так, как современные головастики от лягушек. Кладка яиц на суше способствовала образованию у них нескольких оболочек. Оболочки предохраняли яйца от механического повреждения и высыхания, обеспечивали зародыш воздухом. Волокнистая и известковая оболочки предохраняли яйца от растекания, механических повреждений и проникновения бактерий. Белковая оболочка содержала основные запасы воды. Часть ее выделялась в результате окисления жиров, часть поступала из наружной оболочки. По мере развития зародыша возникали и другие оболочки.
Эволюция пресмыкающихся происходила очень быстро, поскольку на суше еще не было животных, способных конкурировать с ними. Еще задолго до конца пермского периода пресмыкающиеся вытесняют стегоцефалов. Примитивные пресмыкающиеся – котилозавры – дали многочисленных потомков, впоследствии захвативших воду, сушу и воздух. Величиной от лягушки до бегемота, они еще имели много признаков стегоцефалов, в частности зубы и ребра, располагавшиеся от шеи до хвоста, короткие массивные конечности. Но строение черепа, позвонков, кожи уже было таким же, как у пресмыкающихся.
Более сложной по сравнению с другими пресмыкающимися организацией обладали парейазавры, размеры которых достигали 3 м. Однако и они имели в плечевом поясе кожные кости, свойственные рыбам и земноводным. Череп парейазавров представлял собой сплошную костную коробку с отверстиями для глаз, ноздрей и теменного органа. Они были травоядными и жили на берегах рек и озер.
В середине пермского периода достигли расцвета котилозавры. В начале триаса они вымерли. Появились более организованные и специализированные пресмыкающиеся – потомки котилозавров. Пермские пресмыкающиеся приспосабливались к разнообразнейшим условиям жизни. Большинство групп животных стали более подвижными, а их скелеты – более легкими. Они питались разнообразной пищей: растениями, моллюсками, рыбой.
Появляются и настоящие хищники – пеликозавры, на позвоночниках которых имелись высокие гребни. У некоторых пресмыкающихся конечности удлиняются, исчезают кожные кости. В височной области черепа возникают дуги, к которым прикреплялась сложная система мышц. Зубы травоядных становятся плоскими, а такой четырехметровый хищник, как иностранцевия, имел уже настоящие клыки.
Среди хищных пресмыкающихся появляются формы, похожие на современных волков, гиен, куниц. Это говорит о том, что образ жизни животных того времени и нынешних был сходным.
Среди пермских пресмыкающихся жили и зверозубые. Их зубы, как и зубы млекопитающих, различались между собой по форме. У них были резцы, клыки и бугристые коренные зубы. Нижняя челюсть состояла из одной зубной кости, а не из нескольких, как у рыб, земноводных и типичных пресмыкающихся. Подобно млекопитающим, зверозубые пресмыкающиеся имели вторичное костное нёбо, отделявшее носоглотку от ротовой полости. Это позволяло млекопитающим жевать пищу. Зверозубые похожи на млекопитающих также по строению лопаток и таза. Все это свидетельствует о том, что зверозубые были предками млекопитающих.
Пресмыкающиеся разных и одних и тех же пермских материков существенно отличались друг от друга. Очевидно, в пермский период существовали различные климатические зоны.
С пермским периодом связаны значительные месторождения угля, каменной и калийной солей, железной руды, меди, цветных металлов, ртути и нефти.
Продолжался пермский период 55 млн. лет.
Мезозойская эра
Мезозойская эра – это эра средней жизни. Она названа так потому, что флора и фауна данной эры являются переходными между палеозойской и кайнозойской. В мезозойскую эру постепенно формируются современные очертания материков и океанов, современная морская фауна и флора. Образовались Анды и Кордильеры, горные массивы Китая и Восточной Азии. Сформировались впадины Атлантического и Индийского океанов. Началось формирование впадин Тихого океана.
Подразделяется мезозойская эра на три периода: триасовый, юрский и меловой.
Триасовый период
Триасовый период получил свое название от того, что к его отложениям причисляют три разных комплекса пород: нижний – континентальный песчаник, средний – известняковый и верхний – нейпер.
Наиболее характерными отложениями триасового периода являются: континентальные песчано‑глинистые породы (нередко с линзами угля); морские известняки, глины, сланцы; лагунные ангидриты, соли, гипсы.
В триасовый период северный материк Лавразия соединился с южным – Гондваной. Большой залив, начинавшийся на востоке Гондваны, простирался вплоть до северного побережья современной Африки, затем поворачивал на юг, почти полностью отделяя Африку от Гондваны. С запада тянулся длинный залив, отделявший западную часть Гондваны от Лавразии. На Гондване возникло много впадин, постепенно заполнявшихся континентальными отложениями.
В среднем триасе усилилась деятельность вулканов. Обмелевают внутриконтинентальные моря, образуются многочисленные впадины. Начинается формирование горных хребтов Южного Китая и Индонезии. На территории современного Средиземноморья климат был теплым и влажным. Более прохладным и влажным он был в тихоокеанской зоне. На территории Гондваны и Лавразии господствовали пустыни. Холодным и сухим был климат северной половины Лавразии.
Наряду с изменениями в распределении моря и суши, формированием новых горных массивов и вулканических областей, интенсивно проходила смена одних животных и растительных форм другими. Лишь немногие семейства перешли из палеозойской эры в мезозойскую. Это дало основание некоторым исследователям для утверждения о великих катастрофах, происшедших на рубеже палеозоя и мезозоя. Однако при исследовании отложений триасового периода можно легко убедиться, что резкой грани между ними и пермскими отложениями не существует, следовательно, одни формы растений и животных сменялись другими, вероятно, постепенно. Главной причиной являлись не катастрофы, а эволюционный процесс: более совершенные формы постепенно вытесняли менее совершенные.
Сезонное изменение температур триасового периода начало оказывать заметное влияние на растения и животных. Отдельные группы пресмыкающихся приспособились к холодным сезонам. Именно от этих групп в триасе произошли млекопитающие, а несколько позднее – и птицы. В конце мезозойской эры климат стал еще более холодным. Появляются листопадные древесные растения, которые в холодные сезоны частично или полностью сбрасывают листья. Данная особенность растений – это приспособление к более холодному климату.
Похолодание в триасовом периоде было незначительным. Сильнее всего оно проявилось в северных широтах. На остальной территории было тепло. Поэтому пресмыкающиеся чувствовали себя в триасовом периоде достаточно хорошо. Разнообразнейшие их формы, с которыми мелкие млекопитающие еще не способны были конкурировать, расселились по всей поверхности Земли. Необычайному расцвету пресмыкающихся способствовала также богатая растительность триасового периода.
В морях развились исполинские формы головоногих моллюсков. Диаметр раковин некоторых из них был до 5 м. Правда, и теперь в морях обитают исполинские головоногие моллюски, например кальмары, достигающие 18 м в длину, однако в мезозойскую эру исполинских форм было намного больше.
Состав атмосферы триасового периода по сравнению с пермским изменился мало. Климат стал более влажным, однако пустыни в центре континента оставались. Некоторые растения и животные триасового периода дожили до наших дней в районе Средней Африки и Южной Азии. Это говорит о том, что состав атмосферы и климат отдельных участков суши почти не изменились на протяжении мезозойской и кайнозойской эр.
И все же стегоцефалы вымерли. Их вытеснили пресмыкающиеся. Более совершенные, подвижные, хорошо приспособленные к разнообразным условиям жизни, они питались той же пищей, что и стегоцефалы, селились в тех же местах, поедали молодняк стегоцефалов и в конечном итоге истребили их.
Среди триасовой флоры изредка еще встречались каламиты, семенные папоротники и кордаиты. Преобладали настоящие папоротники, гинкговые, беннетитовые, саговниковые, хвойные. Саговниковые существуют и в настоящее время в районе Малайского архипелага. Они известны под названием саговых пальм. По своему внешнему виду саговниковые занимают промежуточное место между пальмами и папоротниками. Ствол саговниковых довольно толстый, колоннообразный. Крона состоит из жестких перистых листьев, расположенных венчиком. Размножаются растения при помощи макро‑ и микроспор.
Папоротники триаса представляли собой прибрежные травянистые растения, имевшие широкие рассеченные листья с сетчатым жилкованием. Из хвойных растений хорошо изучена вольтция. Она имела густую крону и такие, как у ели, шишки.
Гинкговые были довольно высокими деревьями, их листья образовывали густые кроны.
Особое место среди триасовых голосеменных занимали беннетиты – деревья с мутовчатыми крупными сложными листьями, напоминающими листья саговников. Органы размножения беннетитов занимают промежуточное место между шишками саговников и цветками некоторых цветковых растений, в частности магнолиевых. Таким образом, вероятно, именно беннетитов следует считать предками цветковых растений.
Из беспозвоночных триасового периода известны уже все типы животных, существующих и в наше время. Наиболее характерными морскими беспозвоночными были рифообразующие животные и аммониты.
В палеозое уже существовали животные, которые колониями покрывали дно моря, образуя рифы, хотя и не очень мощные. В триасовый период, когда вместо табулят появляется много колониальных шестилучевых кораллов, начинается формирование рифов толщиной до тысячи метров. Чашечки шестилучевых кораллов имели шесть или двенадцать известковых перегородок. В результате массового развития и быстрого роста кораллов на дне моря образовывались подводные леса, в которых селились многочисленные представители других групп организмов. Некоторые из них принимали участие в рифообразовании. Двустворчатые моллюски, водоросли, морские ежи, морские звезды, губки жили между кораллами. Разрушаемые волнами, они образовывали грубозернистый или мелкозернистый песок, заполнявший все пустоты кораллов. Вымытый волнами из этих пустот, известковый ил откладывался в бухтах и лагунах.
Довольно характерны для триасового периода некоторые двустворчатые моллюски. Их тонкие, как бумага, раковины с хрупкими ребрами образуют в отдельных случаях целые прослойки в отложениях данного периода. Жили двустворчатые моллюски в мелководных илистых бухтах – лагунах, на рифах и между ними. В верхнетриасовом периоде появляется множество толстораковинных двустворчатых моллюсков, прочно прикреплявшихся к известняковым отложениям мелководных бассейнов.
В конце триаса в связи с усилением вулканической деятельности часть известняковых отложений покрылась пеплом и лавами. Поднимавшийся из недр Земли пар принес с собой многие соединения, из которых образовались месторождения цветных металлов.
Самыми распространенными из брюхоногих моллюсков были переднежаберные. Широко распространились в морях триасового периода аммониты, раковины которых в отдельных местах скопились в огромном количестве. Появившись в силурийском периоде, они на протяжении всей палеозойской эры еще не играли большой роли среди других беспозвоночных. Аммониты не могли успешно конкурировать с довольно сложными наутилоидеями. Раковины аммонитов были образованы из известковых пластинок, имевших толщину папиросной бумаги и потому почти не защищавших мягкое тело моллюска. Только когда их перегородки согнулись в многочисленные складки, раковины аммонитов приобрели прочность и превратились в настоящее укрытие от хищников. С усложнением перегородок раковины сделались еще более прочными, а наружное строение дало им возможность приспособиться к самым разнообразным условиям жизни.
Представителями иглокожих были морские ежи, лилии и звезды. На верхнем конце тела морских лилий располагалась похожая на цветок основная часть. В ней различают венчик и хватательные органы – «руки». Между «руками» в венчике находились ротовое и анальное отверстия. «Руками» морская лилия загребала в ротовое отверстие воду, а вместе с ней и морских животных, которыми питалась. Стебель многих триасовых морских лилий был спиральным.
В триасовых морях обитали известковые губки, мшанки, листоногие раки, остракоды.
Рыбы были представлены водившимися в пресных водоемах акулами и населявшими море моллюскоидами. Появляются первые примитивные костистые рыбы. Мощные плавники, хорошо развитый зубной аппарат, совершенная форма, прочный и легкий скелет – все это способствовало быстрому распространению костистых рыб в морях нашей планеты.
Земноводные были представлены стегоцефалами из группы лабиринтодонтов. Это были малоподвижные животные с небольшим туловищем, маленькими конечностями и крупной головой. Они лежали в воде в ожидании добычи, и когда добыча приближалась, хватали ее. Их зубы имели сложную лабиринтообразную складчатую эмаль, поэтому их и назвали лабиринтодонтами. Кожа увлажнялась слизистыми железами. Другие амфибии выходили на сушу охотиться на насекомых. Наиболее характерные представители лабиринтодонтов – мастодонозавры. Эти животные, черепа которых достигали одного метра в длину, по внешнему виду напоминали огромных лягушек. Они охотились на рыб и поэтому редко оставляли водную среду.
Мастодонозавр.
Болот становилось меньше, и мастодонозавры вынуждены были заселять все более глубокие места, часто скапливаясь в большом количестве. Вот почему теперь находят много их скелетов на небольших участках.
Пресмыкающиеся в триасе характеризуются значительным разнообразием. Появляются новые группы. Из котилозавров остаются лишь проколофоны – небольшие животные, питавшиеся насекомыми. Чрезвычайно любопытную группу пресмыкающихся представляли архозавры, к которым принадлежали текодонты, крокодилы и динозавры. Представители текодонтов, величиной от нескольких сантиметров до 6 м, являлись хищниками. Они еще отличались рядом примитивных особенностей и походили на пермских пеликозавров. Одни из них – псевдозухии – имели длинные конечности, длинный хвост и вели наземный образ жизни. Другие, в том числе крокодилообразные фитозавры, обитали в воде.
Крокодилы триасового периода – небольшие примитивные животные протозухии – жили в пресных водоемах.
Среди динозавров появляются тероподы и прозауроподы. Тероподы передвигались на хорошо развитых задних конечностях, имели тяжелый хвост, мощные челюсти, небольшие и слабые передние конечности. По величине эти животные были от нескольких сантиметров до 15 м. Все они относились к хищникам.
Прозауроподы питались, как правило, растениями. Некоторые из них были всеядными животными. Ходили они на четырех ногах. Прозауроподы имели небольшую голову, длинную шею и хвост.
Представители подкласса синаптозавров вели самый разнообразный образ жизни. Трилофозавр лазил по деревьям, питался растительной пищей. По внешнему виду он напоминал кошку.
Тюленеобразные пресмыкающиеся жили у побережья, питались главным образом моллюсками. Плезиозавры обитали в море, но иногда выходили на берег. Они достигали 15 м в длину. Питались рыбой.
В отдельных местах довольно часто находят отпечатки ступней огромного животного, ходившего на четырех ногах. Его назвали хиротериумом. По сохранившимся отпечаткам можно представить строение ступни этого животного. Четыре неуклюжих пальца окружали толстую мясистую подошву. На трех из них были когти. Передние конечности хиротериума почти втрое меньше задних. На влажном песке животное оставляло глубокие следы. При отложении новых пластов следы постепенно окаменевали. Позднее суша была залита морем, скрывшим следы. Они оказались засыпанными морскими отложениями. Следовательно, в ту эпоху неоднократно заливало море. Острова опускались ниже уровня моря, и жившие на них животные вынуждены были приспосабливаться к новым условиям. В море появляется много пресмыкающихся, которые, несомненно, произошли от материковых предков. Быстро развились черепахи с широким костяным панцирем, дельфинообразные ихтиозавры – рыбоящеры и исполинские плезиозавры с маленькой головой на длинной шее. Преобразуются их позвонки, изменяются конечности. Шейные позвонки ихтиозавра срастаются в одну кость, а у черепах разрастаются, образуя верхнюю часть панциря.
Ихтиозавр имел ряд однородных зубов, у черепах зубы исчезают. Пятипалые конечности ихтиозавров превращаются в хорошо приспособленные для плавания ласты, у которых трудно различить плечо, предплечье, запястье и кости пальцев.
Начиная с триасового периода, пресмыкающиеся, перешедшие жить в море, постепенно заселяют все более обширные пространства океана.
Древнейшее млекопитающее, найденное в триасовых отложениях Северной Каролины, называется дроматериумом, что в переводе значит «бегающий зверь». Этот «зверь» был всего 12 см в длину. Дроматериум принадлежал к яйцекладущим млекопитающим. Они, подобно современным австралийской ехидне и утконосу, не рождали детенышей, а откладывали яйца, из которых вылупливались недоразвитые детеныши. В отличие от пресмыкающихся, совершенно не заботившихся о своих потомках, дроматериумы выкармливали детенышей молоком.
С отложениями триасового периода связаны месторождения нефти, природных газов, бурого и каменного угля, железной и медной руд, каменной соли.
Длился триасовый период 35 млн. лет.
Юрский период
Впервые отложения данного периода были найдены в Юрах (горы в Швейцарии и Франции), отсюда и произошло название периода. Юрский период подразделяется на три отдела: лейяс, догер и мальм.
Отложения юрского периода довольно разнообразны: известняки, обломочные породы, сланцы, магматические породы, глины, пески, конгломераты, сформировавшиеся в разнообразнейших условиях.
Значительно распространены осадочные породы, содержащие множество представителей фауны и флоры.
Интенсивные тектонические движения в конце триасового и в начале юрского периодов способствовали углублению больших заливов, постепенно отделивших Африку и Австралию от Гондваны. Углубился залив между Африкой и Америкой. В Лавразии сформировались впадины: Немецкая, Англо‑Парижская, Западно‑Сибирская. Арктическое море залило северное побережье Лавразии.
Интенсивный вулканизм и горообразовательные процессы обусловили формирование Верхоянской складчатой системы. Продолжалось формирование Анд и Кордильер. Теплые морские течения достигли арктических широт. Климат стал теплым и влажным. Об этом говорит значительное распространение коралловых известняков и остатки теплолюбивой фауны и флоры. Совсем мало встречается отложений сухого климата: лагунных гипсов, ангидритов, солей и красных песчаников. Уже существовало холодное время года, однако оно характеризовалось лишь понижением температуры. Не было ни снега, ни льда.
Климат юрского периода зависел не только от солнечного света. Множество вулканов, излияния магмы на дно океанов подогревали воду и атмосферу, насыщали воздух паром воды, выпадавшей затем дождями на сушу, бурными потоками стекавшей в озера и океаны. Об этом свидетельствуют многочисленные пресноводные отложения: белые песчаники, чередующиеся с темными суглинками.
Теплый и влажный климат благоприятствовал расцвету растительного мира. Папоротникообразные, цикадовые, хвойные образовывали обширные болотистые леса. На побережье произрастали араукарии, туи, цикадовые. Папоротники и хвощи образовывали подлесок. В нижней юре на всей территории северного полушария растительность была довольно однообразной. Но уже начиная со средней юры, можно определить два растительных пояса: северный, в котором преобладали гинкго и травянистые папоротники, и южный с беннетитами, цикадовыми, араукариями, древовидными папоротниками.
Характерными папоротниками юрского периода являлись матонии, сохранившиеся до сих пор на Малайском архипелаге. Хвощи и плауны почти не отличались от современных. Место вымерших семенных папоротников и кордаитов занимают саговники, растущие и теперь в тропических лесах.
Значительно распространены были также гинкговые. Их листья обращались к солнцу ребром и напоминали огромные веера. От Северной Америки и Новой Зеландии до Азии и Европы росли густые леса хвойных растений – араукарий и беннетитов. Появляются первые кипарисовые и, возможно, еловые.
К представителям юрских хвойных относится также секвойя – современная гигантская калифорнийская сосна. В настоящее время секвойи остались только на Тихоокеанском побережье Северной Америки. Сохранились отдельные формы еще более древних растений, например глассоптерис. Но таких растений немного, поскольку они были вытеснены более совершенными.
Буйная растительность юрского периода способствовала широкому распространению пресмыкающихся. Значительно развились динозавры. Среди них выделяют ящеротазовых и птицетазовых. Ящеротазовые передвигались на четырех ногах, имели на ступнях по пять пальцев, питались растениями. Большинство из них имели длинную шею, маленькую голову и длинный хвост. У них было два мозга: один небольшой – в голове; второй значительно больший по величине – у основания хвоста.
Самым крупным из юрских динозавров был брахиозавр, достигавший в длину 26 м, весивший около 50 т. У него были столбовидные ноги, маленькая голова, толстая длинная шея. Жили брахиозавры на берегах юрских озер, питались водной растительностью. Ежедневно брахиозавру было нужно не менее полутонны зеленой массы.

Брахиозавр.
Диплодок – древнейшее пресмыкающееся, его длина составляла 28 м. У него была длинная тонкая шея и длинный толстый хвост. Подобно брахиозавру, диплодок передвигался на четырех ногах, задние были длиннее передних. Бóльшую часть своей жизни диплодок проводил на болотах и озерах, где пасся и спасался от хищников.

Диплодок.
Бронтозавр был сравнительно высоким, имел большой горб на спине и толстый хвост. Длина его составляла 18 м. Позвонки бронтозавра были полые. Долотовидные маленькие зубы густо располагались на челюстях небольшой головы. Обитал бронтозавр в болотах, на берегах озер.

Бронтозавр.
Птицетазовые динозавры подразделяются на двуногих и четвероногих. Разные по величине и внешнему виду, они питались преимущественно растительностью, но среди них уже появляются и хищники.
К растительноядным относятся стегозавры. У них имелось по два ряда больших пластин на спине и парные шипы на хвосте, защищавшие их от хищников. Появляется множество чешуйчатых лепидозавров – мелких хищников с клювовидными челюстями.
В юрский период впервые появляются летающие ящеры. Летали они при помощи кожистой оболочки, натянутой между длинным пальцем кисти и костями предплечья. Летающие ящеры были хорошо приспособлены к полету. Они имели легкие трубкообразные кости. Чрезвычайно удлиненный внешний пятый палец передних конечностей состоял из четырех суставов. Первый палец имел вид маленькой кости либо совсем отсутствовал. Второй, третий и четвертый пальцы состояли из двух, реже трех костей и имели когти. Задние конечности были довольно сильно развиты. На их концах имелись острые когти. Череп летающих ящеров был сравнительно крупным, как правило, удлиненным и заостренным. У старых ящеров черепные кости срастались и черепа становились похожими на черепа птиц. Межчелюстная кость иногда разрасталась в удлиненный беззубый клюв. У зубатых ящеров зубы были простыми и сидели в углублениях. Самые крупные зубы находились спереди. Иногда они торчали в сторону. Это помогало ящерам ловить и удерживать добычу. Позвоночник животных состоял из 8 шейных, 10–15 спинных, 4–10 крестцовых и 10–40 хвостовых позвонков. Грудная клетка была широкой и имела высокий киль. Лопатки были длинными, тазовые кости срослись. Наиболее характерные представители летающих ящеров – птеродактиль и рамфоринх.

Птеродактиль.
Птеродактили в большинстве случаев были бесхвостыми, разными по величине – от размеров воробья до вороны. Они имели широкие крылья и вытянутый вперед узкий череп с небольшим числом зубов в передней части. Птеродактили жили большими стаями на берегах лагун позднеюрского моря. Днем они охотились, а с наступлением ночи укрывались на деревьях или в скалах. Кожа птеродактилей была морщинистой и голой. Питались они главным образом рыбой, иногда морскими лилиями, моллюсками, насекомыми. Для того, чтобы взлететь, птеродактили вынуждены были спрыгивать со скал или деревьев.
Рамфоринхи имели длинные хвосты, длинные узкие крылья, большой череп с многочисленными зубами. Длинные зубы разной величины выгибались вперед. Хвост ящера заканчивался лопастью, служившей рулем. Рамфоринхи могли взлетать с земли. Они селились на берегах рек, озер и морей, питались насекомыми и рыбой.

Рамфоринх.
Летающие ящеры жили только в мезозойскую эру, причем их расцвет приходится на позднеюрский период. Их предками являлись, по‑видимому, вымершие древние пресмыкающиеся псевдозухии. Длиннохвостые формы появились раньше короткохвостых. В конце юрского периода они вымерли.
Следует заметить, что летающие ящеры не были предками птиц и летучих мышей. Летающие ящеры, птицы и летучие мыши произошли и развивались каждый своим путем, и между ними отсутствуют близкие родственные связи. Единственный общий признак для них – умение летать. И хотя все они приобрели эту способность благодаря изменению передних конечностей, отличия в строении их крыльев убеждают нас в том, что у них были совершенно разные предки.
Моря юрского периода населяли дельфинообразные пресмыкающиеся – ихтиозавры. Они имели длинную голову, острые зубы, большие глаза, окруженные костным кольцом. Длина черепа некоторых из них составляла 3 м, а длина тела – 12 м. Конечности ихтиозавров состояли из костных пластинок. Локоть, плюсна, кисть и пальцы по форме мало отличались друг от друга. Около ста костных пластинок поддерживали широкий ласт. Плечевой и тазовый пояса были слабо развиты. На теле имелось несколько плавников. Ихтиозавры были живородящими животными. Наряду с ихтиозаврами жили плезиозавры. У них были толстое туловище с четырьмя ластообразными конечностями, длинная змеевидная шея с маленькой головой.
В юрский период появляются новые роды ископаемых черепах, а в конце периода – и современные черепахи.
Бесхвостые лягушкообразные земноводные обитали в пресных водоемах. В юрских морях было очень много рыб: костистых, скатов, акул, хрящевых, ганоидных. Они имели внутренний скелет из гибкой хрящевой ткани, пропитанной солями кальция: плотный костный чешуйчатый покров, хорошо защищавший их от врагов, и челюсти с крепкими зубами.
Из беспозвоночных в юрских морях водились аммониты, белемниты, морские лилии. Однако в юрский период аммонитов насчитывалось гораздо меньше, чем в триасе. Юрские аммониты отличаются от триасовых и по своему строению, за исключением филоцераса, совершенно не изменившегося при переходе из триаса в юру. Отдельные группы аммонитов сохранили до нашего времени перламутр. Одни животные обитали в открытом море, другие – заселяли заливы и мелкие внутриконтинентальные моря.
Головоногие моллюски – белемниты – целыми стаями плавали в юрских морях. Наряду с небольшими экземплярами были настоящие великаны – длиной до 3 м.
Остатки внутренних раковин белемнитов, известные под названием «чертовы пальцы», встречаются в отложениях юрского периода.
В морях юрского периода получили значительное развитие также двустворчатые моллюски, особенно принадлежащие к семейству устриц. Они начинают образовывать устричные банки.
Значительные изменения претерпевают морские ежи, селившиеся на рифах. Наряду с дожившими до наших дней круглыми формами жили двусторонне‑симметричные неправильной формы ежи. Их тело было вытянуто в одном направлении. Некоторые из них обладали челюстным аппаратом.
Юрские моря были относительно мелководными. Реки приносили в них мутную воду, задерживая газообмен. Глубокие бухты наполнялись гниющими остатками и илом, содержащим большое количество сероводорода. Именно поэтому в подобных местах хорошо сохранились остатки животных, занесенные морскими течениями или волнами.
Губки, морские звезды, морские лилии нередко переполняют юрские отложения. Значительное распространение получили в юрский период «пятирукие» морские лилии. Появляется много ракообразных: усоногие, десятиногие, листоногие раки, пресноводные губки, среди насекомых – стрекозы, жуки, цикады, клопы.
В юрский период появляются и первые птицы. Их предками являлись древние пресмыкающиеся псевдозухии, давшие начало также динозаврам и крокодилам. Наиболее похожа на птиц орнитозухия. Она, подобно птицам, передвигалась на задних ногах, имела прочный таз и была покрыта похожей на перья чешуей. Часть псевдозухий перешла жить на деревья. Их передние конечности специализировались для обхвата пальцами ветвей. На черепе псевдозухии имелись боковые впадины, что значительно уменьшало массу головы. Лазанье по деревьям и прыгание по ветвям укрепили задние конечности. Постепенно расширявшиеся передние конечности поддерживали животных в воздухе и позволяли им планировать. В качестве примера подобного пресмыкающегося можно указать склеромохлюза. Его длинные тонкие ноги свидетельствуют о том, что он хорошо прыгал. Удлиненные предплечья помогали животным лазить и цепляться за ветки деревьев и кустов. Важнейшим моментом в процессе превращения пресмыкающихся в птиц было преобразование чешуи в перья. Сердце животных имело четыре камеры, чем обеспечивалась постоянная температура тела.
В позднеюрский период появляются первые птицы – археоптериксы, величиной с голубя. Кроме коротких перьев, на крыльях у археоптериксов было по семнадцать маховых перьев. Рулевые перья хвоста располагались на всех хвостовых позвонках и были направлены назад и вниз. Одни исследователи считают, что перья птицы были яркими, как у современных тропических птиц, другие – что перья были серого или коричневого цвета, третьи – что они были пестрыми. Масса птицы достигала 200 г. Многие признаки археоптерикса говорят о его родственных связях с пресмыкающимися: три свободных пальца на крыльях, покрытая чешуей голова, прочные конические зубы, состоявший из 20 позвонков хвост. Позвонки птицы были двояковогнутые, как у рыб. Археоптериксы жили в араукариевых и цикадовых лесах. Питались преимущественно насекомыми и семенами.

Археоптерикс.
Среди млекопитающих появились хищники. Небольшие по размерам, они обитали в лесах и густых кустарниках, охотясь на мелких ящеров и других млекопитающих. Некоторые из них приспособились к жизни на деревьях.
С юрскими отложениями связаны месторождения угля, гипса, нефти, соли, никеля и кобальта.
Продолжался данный период 55 млн. лет.
Меловой период
Меловой период получил такое название потому, что с ним связаны мощные отложения мела. Он подразделяется на два отдела: нижний и верхний.
Горообразовательные процессы в конце юрского периода значительно изменили очертания материков и океанов. Северная Америка, отделенная прежде от огромного Азиатского материка широким проливом, соединилась с Европой. На востоке Азия сомкнулась с Америкой. Южная Америка полностью отделилась от Африки. Австралия находилась там же, где она находится и в настоящее время, однако была меньше по размерам. Продолжается формирование Анд и Кордильер, а также отдельных хребтов Дальнего Востока.
В верхнемеловой период море залило обширные площади северных материков. Под водой находились Западная Сибирь и Восточная Европа, бóльшая часть Канады и Аравия. Накопляются мощные толщи мела, песков, мергелей.
В конце мелового периода вновь активизируются горообразовательные процессы, в результате которых сформировались горные хребты Сибири, Анды, Кордильеры и горные хребты Монголии.
Изменился климат. В высоких широтах на севере в меловой период уже была настоящая зима со снегом. В границах современной умеренной зоны часть пород деревьев (орех, ясень, бук) ничем не отличались от современных. Листья этих деревьев опадали на зиму. Однако, как и прежде, климат в целом был гораздо теплее современного. Все еще были распространены папоротники, саговники, гинкговые, беннетиты, хвойные, в частности секвойи, тисы, сосны, кипарисы, ели.
В середине мелового периода буйно развиваются цветковые растения. В то же время они вытесняют представителей древнейшей флоры – споровые и голосеменные растения. Считается, что цветковые растения возникли и развились в северных районах, впоследствии они расселились по всей планете. Цветковые растения намного моложе хвойных, известных нам еще с карбонового периода. Густые леса из гигантских древовидных папоротников и хвощей не имели цветков. Они неплохо приспособились к условиям жизни того времени. Однако постепенно влажный воздух первичных лесов делался все более сухим. Дождей выпадало совсем мало, невыносимо жгло солнце. Высыхала почва в районах первичных болот. На южных материках возникли пустыни. Растения переместились в районы с более прохладным и влажным климатом севера. А затем снова пошли дожди, насыщавшие влажную почву. Климат древней Европы стал тропическим, на ее территории возникли леса, подобные современным джунглям. Вновь отступает море, и растения, населявшие побережье во влажном климате, оказались в более сухом климате. Многие из них погибли, но некоторые приспособились к новым условиям жизни, образовав плоды, защищавшие семена от высыхания. Потомки подобных растений постепенно заселили всю планету.
Почва также претерпевала изменения. Ил, остатки растений и животных обогащали ее питательными веществами.
В первичных лесах пыльца растений разносилась лишь ветром и водой. Однако появились первые растения, пыльцой которых питались насекомые. Часть пыльцы приставала к крыльям и ногам насекомых, и они переносили ее с цветка на цветок, опыляя растения. У опыленных растений семена созревали. Растения же, на которых не побывали насекомые, не размножались. Поэтому распространялись только растения с душистыми цветками разнообразнейших форм и красок.
С появлением цветков изменились и насекомые. Среди них появляются насекомые, которые вообще не могут жить без цветков: бабочки, пчелы. Из опыленных цветков развивались плоды с семенами. Птицы и млекопитающие поедали эти плоды и разносили семена на большие расстояния, распространяя растения на новые участки материков. Появилось много травянистых растений, заселивших степи и луга. Листья деревьев осенью осыпались, а в летнюю жару свертывались.
Растения распространились по Гренландии и островам Северного Ледовитого океана, где было относительно тепло. В конце мелового периода с похолоданием климата появилось много холодостойких растений: ива, тополь, береза, дуб, калина, свойственные и флоре нашего времени.
С развитием цветковых растений к концу мелового периода вымерли беннетиты, значительно уменьшилось количество саговников, гинкговых, папоротников. Наряду с изменением растительности изменялась и фауна.
Значительно распространились фораминиферы, раковины которых образовали мощные отложения мела. Появляются первые нуммулиты. Кораллы образовывали рифы.
Аммониты меловых морей имели раковины своеобразной формы. Если у всех существовавших до мелового периода аммонитов раковины завертывались в одной плоскости, то у меловых аммонитов были вытянутые раковины, согнутые в виде колена, встречались шарообразные и прямые. Поверхность раковин была покрыта шипами.
По мнению одних исследователей, причудливые формы меловых аммонитов являются признаком старения всей группы. Хотя некоторые представители аммонитов еще продолжали размножаться с большой скоростью, их жизненная энергия в меловой период почти иссякла.
По мнению других ученых, аммонитов истребили многочисленные рыбы, ракообразные, рептилии, млекопитающие, а диковинные формы меловых аммонитов являются не признаком старения, а означают попытку каким‑то образом уберечься от прекрасных пловцов, каковыми к тому времени стали костистые рыбы и акулы.
Исчезновению аммонитов способствовало также резкое изменение физико‑географических условий в меловом периоде.
Белемниты, появившиеся намного позднее аммонитов, в меловой период также полностью вымирают. Среди двустворчатых моллюсков были животные, разные по форме и величине, замыкавшие створки с помощью зубчиков и ямок. У устриц и других моллюсков, прикреплявшихся к морскому дну, створки становятся разными. Нижняя створка имела вид глубокой чаши, а верхняя – крышки. У рудистов нижняя створка превратилась в большой толстостенный бокал, внутри которого осталась лишь маленькая камера для самого моллюска. Круглая, похожая на крышку верхняя створка охватывала нижнюю прочными зубами, с помощью которых она могла подниматься и опускаться. Обитали рудисты главным образом в южных морях.
Кроме двустворчатых моллюсков, раковины которых состояли из трех слоев (наружного рогового, призматического и перламутрового), были моллюски с раковинами, имевшими только призматический слой. Это моллюски рода иноцерамус, широко расселившиеся в морях мелового периода, – животные, достигавшие одного метра в диаметре.
В меловой период появляется много новых видов брюхоногих моллюсков. Среди морских ежей особенно возрастает количество неправильных сердцевидных форм. А среди морских лилий появляются разновидности, не имеющие стебля и свободно плавающие в воде с помощью длинных перистых «рук».
Большие изменения произошли и среди рыб. В морях мелового периода постепенно вымирают ганоидные рыбы. Увеличивается количество костистых рыб (многие из них существуют и в наше время). Акулы постепенно приобретают современный вид.
В море еще жили многочисленные пресмыкающиеся. Потомки вымерших в начале мелового периода ихтиозавров достигали 20 м в длину и имели по две пары коротких ластов.
Появляются новые формы плезиозавров и плиозавры. Они жили в открытом море. Крокодилы и черепахи населяли пресноводные и соленоводные бассейны. На территории современной Европы обитали большие ящерицы с длинными шипами на спине и огромные питоны.
Из наземных пресмыкающихся для мелового периода особенно характерными были траходоны и рогатые ящеры. Траходоны могли передвигаться как на двух, так и на четырех ногах. Между пальцами у них имелись перепонки, помогавшие им плавать. Челюсти траходонов напоминали утиный клюв. На них было до двух тысяч мелких зубов.
У трицератопсов были три рога на голове и огромный костяной щит, надежно защищавший животных от хищников. Жили они преимущественно в сухих местах. Питались растительностью.

Трицератопс.
Стиракозавры имели носовые выросты – рога и шесть роговых шипов на заднем крае костяного щита. Их головы достигали двух метров в длину. Шипы и рога делали стиракозавров опасными для многих хищников.
Самым страшным хищным ящером был тиранозавр. Он достигал в длину 14 м. Его череп, длиной более метра, имел большие острые зубы. Передвигался тиранозавр на мощных задних ногах, опираясь на толстый хвост. Его передние ноги были маленькими и слабыми. От тиранозавров остались окаменелые следы, длиной 80 см. Шаг тиранозавра составлял 4 м.

Тиранозавр.
Цератозавр был относительно небольшим, но быстрым хищником. На голове он имел маленький рог, а на спине костяной гребень. Передвигался цератозавр на задних ногах, на каждой из которых было три пальца с большими когтями.
Торбозавр был довольно неповоротливым и охотился главным образом на малоподвижных сколозавров, напоминавших по внешнему виду современных броненосцев. Благодаря мощным челюстям и крепким зубам торбозавры легко разгрызали толстый костяной панцирь сколозавров.

Сколозавр.
Еще продолжали существовать летающие ящеры. Огромный птеранодон, размах крыльев которого составлял 10 м, имел большой череп с длинным костяным гребнем на затылке и длинный беззубый клюв. Тело животного было относительно небольшим. Питались птеранодоны рыбой. Подобно современным альбатросам, они бóльшую часть своей жизни проводили в воздухе. Их колонии находились у моря. Недавно в меловых отложениях Америки были найдены остатки еще одного птеранодона. Размах его крыльев достигал 18 м.

Птеранодон.
Появляются птицы, которые могли хорошо летать. Полностью вымерли археоптериксы. Однако некоторые птицы имели зубы.
У гесперорниса – водоплавающей птицы – длинный палец задних конечностей соединялся с тремя другими короткой плавательной перепонкой. На всех пальцах были когти. От передних конечностей остались только слегка согнутые плечевые кости в виде тонкой палочки. Гесперорнис имел 96 зубов. Молодые зубы росли внутри старых и заменяли их, как только они выпадали. Гесперорнис очень похож на современную гагару. Передвигаться по суше ему было очень трудно.








