 2015-04-30
2015-04-30 3053
3053Головной мозг (лат. encephalon) - передний отдел (центральный орган) центральной нервной системы позвоночных, регулирующий взаимоотношения организма с окружающей средой, управляя поведенческими реакциями и функциями организма. Он состоит из конечного или большого мозга (telencephalon) и мозгового ствола (truncus cerebri). По величине большой мозг превышает все остальные отделы головного мозга вместе взятые (см. главу 15).
Таблица. 31. Некоторые рефлексы, легко наблюдаемые у животных
| Название рефлекса | Раздражение, вызывающее рефлекс | Характер рефлекторного ответа | Центральное звено рефлекторной дуги | |
| Зрачковый Роговичный (корнеальный) Рефлекс холки лошади Коленный рефлекс (у лошади и собаки) Ахиллов рефлекс Копытный рефлекс Сгибательный рефлекс Рефлекс отталкивания Тормозной сухожильный рефлекс Брюшной рефлекс | Внезапное освещение глаза Слабое прикосновение к роговице Легкое дотрагивание до волос или укол кожи в этой области Легкий удар по patella (коленной чашечке) Легкий удар по ахиллову сухожилию Сдавливание копытными щипцами Сгибание в суставе Давление стопы на опору Растяжение сухожилия Проведение штриха по коже животного | Сужение зрачка Смыкание век Сокращение подкожных мышц и вздрагивание кожи Разгибание голени Подошвенное сгибание стопы Вздрагивающее поднятие копыта Отдергивание конечности при повреждении Хромота Расслабление мышцы Сокращение брюшной стенки в области раздражения | Средний мозг Продолговатый мозг Спинной мозг, шейный отдел Спинной мозг, поясничный отдел Спинной мозг, крестцовый отдел Спинной мозг, крестцовый отдел Спинной мозг Спинной мозг Спинной мозг Спинной мозг, грудной отдел | |
В мозговом стволе различают промежуточный мозг (diencephalon), включающий эпиталамус (надбугорье, epithalamus), метаталамус (забугорье, metathalamus), гипоталамус (подбугорье, hуpothalamus), средний мозг (mesencephalon), включающий ножки мозга (pedunculi cerebri) и пластинку крыши (lamina testi); задний мозг (metencephalon), представленный мостом (pons) и мозжечком (cerebellum); продолговатый мозг (mуelencephalon). Задний и продолговатый мозг объединяют под названием ромбовидного мозга (rhombencephalon).
Функции ромбовидного мозга
Выше места выхода корешков первой пары шейных нервов спинномозговой нервный тяж приобретает структуру, характерную для ствола головного мозга. Под названием “ствол” объединяют структуры продолговатого мозга, моста, среднего и отчасти промежуточного мозга (см. выше). В глубине ствола расположена диффузная цепь вставочных нейронов, называемая ретикулярная формация (formatio reticularis). Ретикулярная формация осуществляет активирующее воздействие на кору большого мозга и контролирует рефлекторную деятельность спинного мозга. Ретикулярная формация является местом избирательного действия многих фармакологических средств и ей отводится важная роль в механизме сна и бодрствования, в механизмах формирования целостных, и в частности, условнорефлекторных реакций организма.
В стволе мозга нет четкого сегментарного разграничения серого и белого вещества; черты фрагментарности проявляются в том, что здесь локализованы ядра 12 пар черепномозговых нервов.
Продолговатый мозг и варолиев мост
Белое вещество продолговатого мозга состоит из длинных и коротких волокон. Длинные входят в состав восходящих и нисходящих путей, а короткие осуществляют связи с другими отделами ствола и мозжечком. Серое вещество образует скопления - ядра. Среди них соматические и вегетативные ядра черепномозговых нервов (IX- XII), ядра ретикулярной формации и ядра двигательных путей.
Продолговатый мозг так же как и спинной мозг выполняет две основные функции: рефлекторную и проводниковую.
Таблица 32. Расположение и функции черепномозговых нервов
| Номер пары нервов | Название | Начало афферентных волокон | Окончание эфферентных волокон | Функция | Нерв |
| I II | Обонятельный Зрительный | Обонятельный эпителий носа Сетчатка глаза | - - | Обоняние Зрение | Сенсорный Сенсорный |
| III | Глазодвигательный | Проприорецепторы глазного яблока | Большинство мышц глазного яблока | Мышечное чувство. Движение глаз, их аккомодация и диафрагмирование на поворот глазного яблока | Преимущественно эффекторный. Есть вегетативные волокна |
| IV | Блоковый | Проприорецепторы верхней косой мышцы | Верхняя мышца | Поворот глазного яблока | Преимущественно эффекторный |
| V | Тройничный n.trigeminus | Кожа головы (лица), коньюктива и роговица глаза, зубы, слизистая ротовой полости | Некоторые жевательные мышцы | Кожная рецепция головы (лица) и ротовой полости | Смешанный |
| VI | Отводящий | Проприорецепторы наружной прямой мышцы | Наружная прямая мышца | Поворот глазного яблока к наружи | Преимущественно эффекторный |
| VII | Лицевой | Вкусовые почки передней части языка | Мышцы морды (лица), подчелюстная и подъязычная железы | Вкус, мимика | Преимущественно эффекторный. Есть вегетативные волокна |
| VIII | Слуховой n.acusticus | Улитка, полукружные каналы | - | Слух, положение головы в пространстве и ее движение | Сенсорный |
| IX | Языкогло-точный | Вкусовые почки языка, слизистая глотки | Мышцы глотки, околоушная железа | Глотание, слюноотделение | Смешанный |
| X | Блуждающий | Интерорецепторы органов кровообращения, дыхания, пищеварения | Сердце, желудок, кишечник, печень, почки | Регуляция работы внутренних органов | Смешанный, вегетативный |
| XI | Добавочный | Проприорецепторы мышц плеча | Мышцы плеча | Мышечное чувство, движение плеча | Преимущественно эффекторный |
| XII | Подъязычный | Проприорецепторы мышц языка | Мышцы языка | Мышечное чувство, движение языка | Преимущественно эффекторный |
Механизм рефлекторной деятельности продолговатого мозга принципиально не отличается от подобной функции спинного мозга, т.к. афферентную импульсацию он получает из спинного мозга, а также от рецепторов V, VIII, IX и Х черепномозговых нервов. Эфферентную часть дуг соматических и вегетативных рефлексов продолговатого мозга составляют волокна соответствующих черепномозговых нервов.
В продолговатом мозге находятся следующие нервные центры: дыхания, сердечной деятельности, сосудодвигательный, рефлексов сосания, глотания, слюноотделения, жевания, отделения желудочного и поджелудочного соков, рвоты, кашля, чихания, углеводного обмена и др. Рефлексы, осуществляемые продолговатым мозгом можно разделить на следующие:
1) гемодинамические - регуляция деятельности сердца и просвета сосудов;
2) пищевые - рефлексы сосания, жевания, глотания, отделение пищеварительных соков, сокращение кишечника;
3) защитные - мигание, кашель, чихание, слезоотделение, рвота;
4) дыхательные - сокращение и расслабление межреберных мышц, диафрагмы.
В области продолговатого мозга и варолиева моста лежат ядра следующих черепномозговых нервов: тройничного (V), отводящего (VI), лицевого (VII), слухового (VIII), языкоглоточного (IX), блуждающего (X), добавочного (XI), подъязычного (XII) нервов.
Продолговатый мозг осуществляет через ядра Дейтерса и Бехтерева функцию регуляции мышечного тонуса (рис. 90). За счет мышечного тонуса преодолевается действие силы тяжести, тело сохраняет равновесие. Роль продолговатого мозга в двигательных реакциях животного.
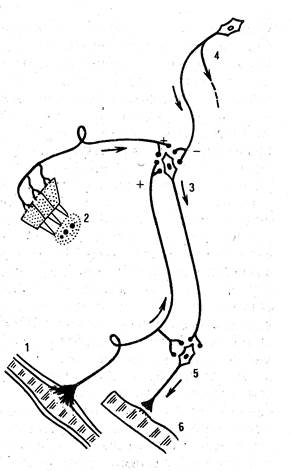
Рис. 90. Центры, контролирующие мышечный тонус:
1- мышечный рецептор; 2- отолитовый орган; 3- ядро Дейтерса; 4- красное ядро; 5- моторная клетка спинного мозга; 6- мышцы; плюс- возбуждение; минус- торможение.
Если головной мозг вместе со средним мозгом и его красными ядрами удален (см. рисунок), но продолговатый мозг сохранен и не отделен от спинного, но у млекопитающих развивается сильное, часами и днями удерживаемое напряжение разгибательных мышц. Конечности такого животного вытянуты и согнуть их можно лишь с трудом. Шейные мышцы, разгибающие голову, напряжены, позвоночник несколько выгнут дугой кверху (опистотонус).
Таким образом, все мышцы, напряжение которых противостоит действию силы тяжести (экстензоры), оказываются после отделения продолговатого мозга от среднего в состоянии длительного тонического напряжения. Это резкое усиление тонуса разгибательных мышц носит название децерибрационной ригидности. Она прекращается после разрушения ядра Дейтерса, а также после отделения его от спинного мозга. Из ядра Дейтерса к мотонейронам спинного мозга поступают импульсы, вызывающие усиление тонуса разгибателей. В ядро Дейтерса из красного ядра непрерывно идут импульсы, снижающие активность его. Возбуждение ядра Дейтерса поддерживается импульсами от рецепторов разгибательных мышц и вестибулярного аппарата, расположенного во внутреннем ухе.
Импульсы, уменьшающие активность ядра Дейтерса идут из красного ядра, мозжечка, подкорковых образований (ганглии) и коры больших полушарий.
Функциональная связь продолговатого мозга с вестибулярным аппаратом и мышечными рецепторами проявляется также в рефлексах позы (положения), которые относятся к тоническим статическим рефлексам.
Проводниковая функция. На нижней поверхности продолговатого мозга находятся два продольных возвышения - пирамиды, которые как бы продолжаются в нисходящие тракты спинного мозга. К наружи от пирамид лежат оливы, от которых начинается оливоспинальный пучок. Волокна этого пучка принимают участие в управлении тоническими рефлексами и непроизвольными локомоторными актами. На дорсальной поверхности простираются дорсальные восходящие тракты (нежный и клиновидный), идущие из спинного мозга до одноименных ядер продолговатого мозга (восходящие пути). Здесь происходит переключение восходящих путей спинного мозга и передача импульсации).
Варолиев мост - мост головного мозга (лат Pons Varolii) располагается на основании мозга, массивный волокнистый тяж, переходящий без ясно видимой границы в средние ножки мозжечка. Латерально из моста отходят тройничные нервы. В мосте расположены ядра черепномозговых нервов, входящие в центры, регулирующие слезотечение, голосообразование, мигание, жевание, глотание. Проводниковая функция заключается в том, что мост связывает вышележащие и нижележащие отделы ЦНС между собой и мозжечком. В нем расположен также центр пневмотаксиса, участвующий в регуляции дыхания.
Средний мозг расположен между варолиевым мостом и промежуточным мозгом.
К основным образованиям среднего мозга относятся: крыша среднего мозга, или пластинка четверохолмия; ножки мозга, мозговой (сильвиев) водопровод, соединяющий III и IV желудочки мозга.
Четверохолмие разделяется на два передних бугра и два задних. Передние бугры являются подкорковыми центрами зрения, задние - подкорковыми центрами слуха. При их участии осуществляется поворот глаз в сторону зрительных и поворот ушей и головы в сторону звуковых раздражителей.
Раздражение в эксперименте четверохолмия электрическим током вызывает те же ориентировочные, настораживающие реакции (вскакивание, поворот ушных раковин, поворот головы, настороженность, движение глаз), которые вызываются в норме сильным и внезапным слуховым или зрительным раздражителем.
Таким образом, пара передних бугров четверохолмия связана с ориентировочными зрительными, а пара задних - с ориентировочными акустическими рефлексами. Ножки мозга включают проводящие пути к промежуточному мозгу (таламусу), базальным ганглиям и коре больших полушарий. С мозжечком средний мозг связан посредством верхних ножек мозжечка, со спинным - тектоспинальным трактом.
Каждая ножка состоит из трех частей: покрышки, черной субстанции и основания. В покрышке находятся красное ядро и ядра блокового и глазодвигательного нервов. Эти нервы идут к мышечному аппарату глазного яблока, обеспечивая координацию движения глаз. Блоковый нерв интернирует верхнюю косую мышцу глаза, а глазодвигательный - верхнюю косую мышцу глаза, а также мышцу, поднимающую веко. Кроме того, в состав глазодвигательного нерва входят парасимпатические волокна, иннервирующие гладкую мускулатуру ресничной мышцы и сфинктер зрачка.
В центральной части среднего мозга находится крупная ретикулярная формация, образующая единый функциональный комплекс с аналогичными структурами спинного, продолговатого и промежуточного мозга. Состоит она из многочисленных, обширно ветвящихся нейронов, отростки которых идут как в восходящем, так и нисходящем направлениях. На ее клетках конвергируют различные афферентные импульсы, что обеспечивает непрерывный, тонический характер ее активности.
Нейроны ретикулярной формации обладают высокой чувствительностью к различным химическим агентам - медиаторам, гормонам, метаболитам, лекарственным веществам. В нижних отделах ствола (продолговатом, среднем и мосту) они являются адренгическими, а в верхних отделах (промежуточный мозг) - холинергическими. На кору головного мозга формация оказывает непосредственное тонизирующее (активирующее) влияние, поэтому ее определяют как неспецифическую активирующую систему.
Клеточные элементы среднего мозга образуют скопления в виде ядер. Среди них важную роль играют черная субстанция и особенно красное ядро.
Черная субстанция принимает участие в осуществлении актов жевания и глотания, в регуляции сложных тонких двигательных актов и эмоционального поведения (через аксоны дофаминергичеких нейронов, идущих в задние отделы лобных долей и центральную извилину коры больших полушарий, а также ряда вегетативных функций (дыхания, тонуса сосудов).
Исключительное значение в красном мозге млекопитающих имеет красное ядро, которое связано с вентральными рогами спинного мозга (начало нисходящего руброспинального тракта).
Красное ядро постоянно посылает тормозящие импульсы в центры продолговатого мозга, ответственные за развитие децеребрационной ригидности. Основная функция красного ядра - регуляция и перераспределение тонуса мышц в зависимости от положения тела в пространстве. Оно осуществляет рефлексы, регулирующие поддержание нормальной позы и равновесия тела: тонические, установочные и статокинетические рефлексы (табл. 4). Тонические и установочные рефлексы обеспечивают принятие основного положения, а также принятие и поддержание постоянной позы (табл. 33).
Таблица 33. Рефлексы, регулирующие положение тела в про-
странстве, которые сохраняются у мезэнцефального животного
| Статические | рефлексы | Статокинетические рефлексы |
| Позотонические (позы) | Выпрямительные (установочные) | |
| Тонические шейные | Лабиринтные выпрямительные | Повороты головы Нистагм (вестибулоглазо-двигательные) |
| Тонические лабиринтные | Шейные выпрямительные | Лифтная реакция |
| Компенсаторная установка глаз | - | Принятие позы при свободном падении, рефлекс наклона |
Как видно из табл. 33, тонические рефлексы делят на 2 большие группы: статические и статокинетические.
Статические рефлексы проявляются при спокойном состоянии, лежании или сидении в различных положениях. Выделяют две группы данных рефлексов: рефлексы положения или позотонические, или позы и выпрямительные, или установочные.
Рефлексы позы. При изменении положения головы раздражаются рецепторы мышц и связок шеи, рецепторы вестибулярного аппарата и перераспределяется тонус мышц конечностей. Изменение положения головы всегда сопровождается не только раздражением нервно-эпителиальных клеток вестибулярного аппарата, но и рецепторы мышц и связок шеи. Поэтому рефлексы позы всегда являются комплексом лабиринтных и шейных рефлексов.
Рефлексы позы можно наблюдать в естественных условиях обитания животного. Например, если животное наклоняет голову, то сгибаются его передние конечности, а задние выпрямляются (при вставании крупного рогатого скота).
Выпрямительные рефлексы способствуют восстановлению нормальной позы тела, если она оказалась нарушенной. Эти рефлексы наиболее выражены у грызунов, у кошки.
Если у животного удалены отделы, лежащие выше среднего мозга, то выпрямительные рефлексы сохраняются. Следовательно, центры данных рефлексов расположены в среднем мозге.
Если мезэнцефальное животное положить на бок, оно поднимает голову и держит ее в естественном положении - теменем кверху. Этот рефлекс начинается с раздражения рецепторов вестибулярного аппарата. Для этого чтобы вестибулярные реакции не осложнились кожными, животное исследуют в воздухе, поддерживая за грудную клетку и таз, минимально касаясь его кожной поверхности. Если придать туловищу любое положение, голова сохраняет нормальную ориентировку - теменем кверху. При разрушении лабиринтов выпрямительный рефлекс на голову животного, подвешенного в воздухе отсутствует, голова повисает, подчиняясь действию силы тяжести. Указанный рефлекс назван рефлексом вестибулярного аппарата на голову.
Но если мезэнцефальное животное без обоих лабиринтов, которое не способно правильно ориентировать свою голову в воздухе, положить на какую-нибудь твердую поверхность - на пол, стол, то вследствие соприкосновения кожи туловища с твердой поверхностью возникает рефлекс, в результате которого голова выпрямляется. Этот рефлекс называют рефлексом с рецепторов кожи туловища на голову.
С помощью двух описанных рефлексов - рефлекса с вестибулярного аппарата на голову и рефлекса с рецепторов кожи туловища на голову - обеспечивается выпрямление головы. Но подъем головы - это только первая фаза выпрямительных рефлексов.
Вторая фаза заключается в рефлекторном выпрямлении туловища, этот рефлекс также начинается с раздражения двух рецепторных полей: проприорецепторов мышц и связок шеи и рецепторов кожи туловища.
Когда в результате первой фазы голова приходит в нормальное положение - теменем кверху, а туловище еще лежит на боку, осуществляется поворот шеи. При этом раздражаются проприорецепторы шейных мышц, что служит исходной точкой для третьего рефлекса - выпрямления туловища. Данный рефлекс назван шейным рефлексом выпрямления или рефлексом с проприорецепторов шеи на выпрямление туловища. Имеется четвертый выпрямительный рефлекс с рецепторов кожи на выпрямление туловища.
У высших млекопитающих (кошка, собака, обезьяна) наблюдается еще одна группа выпрямительных рефлексов, которые начинаются с рецепторов зрительного аппарата. Эти рефлексы получили название оптические выпрямительные рефлексы. Оптические выпрямительные рефлексы осуществляются при обязательном участии коры больших полушарий.
Статокинетические рефлексы возникают у животного при движении: при угловом ускорении (например, при вращении) или линейном ускорении (нарастающем или убывающем), а также при перемещении отдельных частей тела. Центр этих рефлексов - красное ядро среднего мозга. Рецептивным полем всех без исключения статокинетических рефлексов служат рецепторы полукружных каналов внутреннего уха.
Яркий пример статокинетических рефлексов - рефлекс “лифта”: при быстром подъеме конечности сгибаются, а при быстром опускании - разгибаются.
При вращении возникает нистагм головы и глаз, вестибулоглагодвигательные рефлексы, выражающиеся в непроизвольном ритмическом пилообразном движении головы и глаз. В начале голова и глаза медленно поворачиваются в сторону, противоположную направлению вращения, а затем быстро возвращаются в исходное положение. У высокоорганизованных животных развитая кора больших полушарий способна подавлять лабиринтные и проприорецептивные рефлексы. Вместо жесткофиксированных тонических рефлексов у этих животных главное значение имеют изменчивые сложные приобретенные двигательные формы поведения.
Мозжечок (лат. cerebellum) - участвует в координации движения, регуляции мышечного тонуса, сохранение позы и равновесия тела.
Структурно-функционально мозжечок разделяется на три доли: старый мозжечок (archeocerebellum) в котором располагается центр вестибулярных рефлексов (вестибулярный отдел), древний мозжечок (paleocerebellum), представленный передней долей, простой долькой и задней частью тела мозжечка; новый мозжечок (neocerebellum), включающий среднюю часть тела и большую часть полушарий мозжечка. В белом веществе заложены ядра мозжечка: зубчатое, пробковидное, шаровидное и ядро шатра.
Мозжечок имеет три пары ножек: нижние мозжечковые ножки соединяют его с продолговатым мозгом, средние - с мостом мозга, верхние - с крышкой среднего мозга. Сам мозжечок, как надсегментарное образование, не вступает в прямую связь с рецепторами и эффекторами, однако получает обильную информацию. К нему поступают импульсы от рецепторов кожи, мышц, сухожилий через спинномозжечковые пути, а также через ядра продолговатого мозга и ретикулярной формации. Из продолговатого мозга в мозжечок поступают импульсы от вестибулярного аппарата, а из среднего мозга - зрительные и слуховые импульсы.
Вся информация, поступающая в мозжечок, передается клетками Пуркинье, которые оказывают тормозящее влияние на ядра самого мозжечка (через них на нисходящий ретикулоспинальный тракт) и ядро Дейтерса в продолговатом мозгу (начало вестибулоспинального тракта). Следовательно, тормозные влияния мозжечка контролируют большую часть импульсов - команд, поступающих к спинному мозгу по основным нисходящим путям.
Благодаря таким связям и сложному строению мозжечок участвует в координации движений, сохранении позы и равновесия. Это осуществляется путем перераспределения мышечного тонуса, обеспечения правильного напряжения различных групп мышц при каждом двигательном акте, устранения ненужных, лишних движений.
Функция мозжечка изучена хорошо после его удаления. Впервые мозжечок у животных был удален в 1843 г Лучиани. При удалении или нарушении деятельности мозжечка отмечены расстройства в регуляции движения. Лучиани назвал симптомы, которые наступают после удаления мозжечка: атония, астазия, астения и атаксия.
Атония - потеря или ослабление (дистония) мышечного тонуса, наблюдается через несколько дней после операции.
Астазия - потеря способности к слитному тетаническому сокращению; животное не может стоять неподвижно.
Астения - быстрое утомление мышц, вследствие непрерывного движения и затратой большого количества энергии.
Атаксия - плохая координация движений. Походка шаткая, движения размашистые и неловкие при ходьбе.
Нервные клетки мозжечка функционируют непрерывно, что было обнаружено при изучении электрической активности мозжечка. Электрическая активность мозжечка довольна высока и характеризуется высокой частотой (150-250 колебаний в секунду). В мозжечке обнаружены зоны задних конечностей, туловища, передних конечностей и головы, а также слуховая и зрительная зоны.
Раздражение определенных зон (областей) мозжечка вызывает моторные реакции, причем раздражение точек, отстоящих друг от друга всего на 1 мм вызывает различные движения. Это свидетельство огромной роли мозжечка в моторных функциях организма.
Между мозжечком и корой больших полушарий существует тесная взаимосвязь, они оказывают определенное влияние друг на друга. Чем больше развита кора, тем больше развит мозжечок: у приматов он сформирован лучше, чем у низших млекопитающих. Чем выше роль коры в двигательных функциях животного, тем большая роль мозжечка и тем значительнее последствия его удаления или нарушения его функции.
Следовательно, мозжечок помогает коре головного мозга при выполнении животным сложно-координированных уточненных двигательных актов.
Подавляя поток возбуждений, идущих по экстрапирамидной системе, мозжечок как бы “освобождает дорогу” для срочных и точных импульсов пирамидных путей.
Мозжечок играет важную роль в регуляции вегетативных функций. При раздражении мозжечка наблюдаются симпатические и парасимпатические эффекты: расширение зрачков, учащение пульса, повышение артериального давления, сердечной деятельности, изменения деятельности желудочно-кишечного тракта, изменение состава крови, изменение энергетического обмена, нарушение углеводного обмена и др.
Следовательно, в мозжечке лежат центры симпатической и парасимпатической иннервации.
Промежуточный мозг (лат. diencephalon), часть мозгового ствола, которая формируется из оставшейся неразделенной задней части переднего мозга. Включает эпиталамус (epithalamus, надбугорье), метаталамус (metathalamus, забугорье), таламус (thalamus, зрительные бугры), гипоталамус (hуpothalamus, подбугорье).
Эпиталамус включает обонятельный центр и железу внутренней секреции эпифиз.
Метаталамус составляют два образования: латеральное коленчатое тело (corpus geniculatum laterale) и медиальное коленчатое тело (corpus geniculatum mediale). Латеральное коленчатое тело характеризуется слоистостью строения и состоит из шести клеточных слоев: двух крупно- и четырех мелкоклеточных. Периферический зрительный путь заканчивается в мелких клетках, центральный берет начало от крупных клеток. Различные отделы сетчатки проецируются на разные участки латерального коленчатого тела. Медиальное коленчатое тело располагается между верхним холмиком четверохолмия и подушкой зрительного бугра. Ядро его состоит из мелко- и крупноклеточной частей. Клетки имеют многоугольную форму. Афферентной системой медиального коленчатого тела являются волокна боковой петли, которые образуют вокруг него капсулу и постепенно заканчиваются в его клетках. От клеток медиального коленчатого тела начинается центральный слуховой путь, который направляется к коре височной области мозга.
Таламус. Зрительные бугры - основной отдел промежуточного мозга, включающий большое количество ядер (около 40). Таламус воспринимает импульсы всех видов чувствительности и передает их в кору больших полушарий, а также в другие образования цнс.
Выделяют вентральный таламус, в состав которого входят срединные, медиальные, вентро-латеральные и задние ядра, а также дорсальный таламус, включающий переднее, медиальное, дорсальное и другие ядра. Таламус является основным коллектором различного вида чувствительности, идущей прежде всего в кору большого мозга. В переключательных (специфических) таламических ядрах имеет место высокая степень специализации и концентрации афферентных импульсов. Медиальные отделы таламуса играют большую роль в условно-рефлекторной деятельности и поведении животных. Таламус является одним из высших центров болевой чувствительности. Болевые сигналы вызывают очень сильную активацию нейронов неспецифических ядер таламуса.
Установлено, что при некоторых поражениях таламуса появляются мучительные болевые ощущения даже при самом незначительном раздражении (прикосновение к коже, звуковое и световое раздражение). Болевые ощущения очень стойкие и почти не поддаются лечению.
В таламусе осуществляются первоначальный анализ и синтез различных сенсорных импульсов. Этот анализ довольно груб (разница температур, например, только в 10-15ОС и больше). Тонкий анализ проводится в коре головного мозга. В таламусе чувствительные импульсы приобретают эмоциональную окраску (приятные и неприятные ощущения).
Специфические ядра таламуса имеют прямую связь с определенными участками коры головного мозга. Они подразделяются на переключающие и ассоциативные. Переключающие ядра обрабатывают и проводят в сенсорные зоны коры головного мозга зрительные, слуховые, тактильные и интерорецептивные сигналы. Ассоциативные ядра получают импульсы по ответвлениям от переключающих ядер и передают их в ассоциативные зоны коры.
Таламус участвует в регуляции мышечного тонуса. Этот тонус в отличие от контрактильного тонуса продолговатого мозга получил название пластического, так как мышцы становятся чрезвычайно пластичными и животное остается в приданной ему позе. При перерезке афферентных проприоцептивных волокон в соответствующих им мышцах пластический тонус пропадает. Следовательно, пластический тонус представляет генерализованный проприоцептивный рефлекс, центр которого находится в таламусе. Пластический тонус играет большую роль, так как он освобождает организм от необходимости непрерывно удерживать все мышцы в напряженном состоянии и сохранять всегда одну определенную позу. Именно благодаря пластическому тонусу животное может принимать различные позы.
Гипоталамус - одно из древнейших образований в головном мозге - располагается вентральнее зрительных бугров. Оно включает в себя собственно подбугорную область и ряд образований у основания мозга: зрительный перекрест, серый бугор, сосцевидное тело и воронку с отходящим от нее нижним мозговым придатком - гипофизом. Нервные клетки гипоталамуса образуют ядра, которые объединяются в крупные группы: преоптическая, передняя, средняя и задняя (насчитывается в гипоталамусе 32 пары ядер). Ядра гипоталамуса связаны со всеми отделами цнс: эфферентные пути от них идут к зрительным буграм и гипофизу, спускаются к среднему, продолговатому и спинному мозгу, оканчиваясь на клетках, аксоны которых образуют преганглионарные волокна вегетативной нервной системы.
Афферентные импульсы поступают в ядра гипоталамической области главным образом из зрительных бугров, через которые в подбугорье проходят также волокна от обонятельной, премоторной и двигательной зон коры мозга.
Гипоталамус служит промежуточным звеном между зрительными буграми и корой мозга, с одной стороны и вегетативной системой - с другой, так как гипоталамус принимает самое активное участие в регуляции вегетативных функций. Именно в ядрах гипоталамуса осуществляется координация деятельности вегетативной нервной системы, согласование функций эндокринных желез и объединение нейрогуморальных механизмов. В ядрах гипоталамуса происходит переключение электрического импульса на химический (выделение нейросекретов).
Гипоталамус участвует в поддержании постоянной температуры тела. Если у животного разрушить серый бугор, то у него нарушаются процессы теплопродукции и теплоотдачи и животное становится пойкилотермным. При раздражении электрическим током ядер переднего гипоталамуса температура тела животного повышается, а заднего гипоталамуса - понижается.
В области вентромедиальных ядер гипоталамуса расположен центр насыщения. Если разрушить боковые (латеральные ядра), где находится центр голода, то животное полностью отказывается от корма. При голодании в крови уменьшается количество глюкозы, аминокислот, жирных кислот и других веществ. Это приводит к активации соответствующих нейронов и формированию поведенческих реакций, направленных на утоление чувства голода.
Дорсолатерально от супраоптического ядра находится центр жажды, который обеспечивает постоянство водного баланса в организме. При раздражении нейронов этой области усиливается потребление воды (полидипсия), а при разрушении - полный отказ животного от потребления воды (адипсия). Выделяют два механизма жажды, один из которых связан с клеточной дегидратацией и активацией осморецепторов преоптической области гипоталамуса, а другой механизм - со снижением объема внеклеточной жидкости, участием гормональной системы ренин-анггиотензин. Активация последней стимулирует гипоталамический центр жажды.
В преоптической области гипоталамуса расположены нейроны, участвующие в регуляции осмотического давления внутренней среды организма (осморецепторы). Они работают по принципу осмометра. В теле такого нейрона имеется вакуоль, заполненная внутриклеточной жидкостью, осмотическое давление которой служит эталоном: с ним сравнивается осмотическое давление внеклеточной среды, омывающей нейрон. Если осмотическое давление тканевой жидкости и крови увеличивается, вода в силу законов осмоса выходит из вакуоли осморецептора, что приводит к уменьшению размеров вакуоли и тела осморецептороной клетки. Частота нервных импульсов, генерируемых осморецептором, увеличивается, что вызывает усиленную секрецию антидиуретического гормона (АДГ), выделяемого задней долей гипофиза. Этот гормон усиливает реабсорбцию воды из первичной мочи в канальцах и собирательных трубочках почек в кровь, что приводит к понижению осмотического давления крови.
При повышении содержания воды в тканевой жидкости и крови вода всасывается в вакуоль осморецепторной клетки, увеличивая размеры и вакуоли и клетки. Частота возникающих импульсов в осморецепторной клетке снижается, секреция АДГ гипофизом уменьшается, почки выделяют большее количество мочи и организм освобождается от излишней воды.
В гипоталамусе находятся центры, регулирующие различные виды обмена веществ: белкового, жирового, углеводного, водносолевого.
Гипоталамус находится в теснейшей морфологической и функциональной связи с гипофизом. Между ними имеются обширные нервные и сосудистые связи, поэтому их объединяют в единую гипоталамо-гипофизную систему. Нейрогормоны гипоталамуса регулируют выделение “тропных” гормонов гипофизом.
Гипоталамус имеет огромное значение для эмоциональных и поведенческих реакций. При раздражении вентромедиального ядра подбугорья кошки проявляется резко выраженная ярость (агрессивно-оборонительный рефлекс).
Центры гипоталамуса формируют относительно простые акты пищевого и эмоционального поведения - чувство жажды, голода и насыщения, поиск воды и пищи; проявления агрессии, страха, удовольствия или неудовольствия. Определенные центры гипоталамуса участвуют также в регуляции размножения и лактации. Регуляция гомеостаза и нейроэндокринные взаимоотношения в организме осуществляются преимущественно преоптической, передней и средней группами ядер гипоталамуса; задние группы ядер принимают участие в регуляции иммуногенеза.
Несмотря на важную роль многочисленных ядер гипоталамуса в жизнедеятельности организма животного гипоталамус находится под неослабным постоянным контролем вышележащих центров головного мозга, с которыми он связан анатомически и функционально как прямыми нервными связями, так и при помощи ретикулярной формации.
Ретикулярная формация (лат. formacio reticularis) - совокупность структур в цнс, которые характеризуются наличием диффузных скоплений клеток разнообразной формы, окруженных многочисленными волокнами, идущими во всех направлениях, в связи с чем данное образование под микроскопом напоминает сеточку. Это послужило основанием для О. Дейтерса назвать его сетчатым образованием.
Ретикулярная формация осуществляет активирующее воздействие на кору большого мозга и контролирует рефлекторную функцию спинного мозга. Активность ретикулярной формации поддерживается непрерывным притоком афферентных импульсов, поступающих в нее по коллатеральным волокнам от сенсорных проводящих путей.
Важную роль в поддержании активности ретикулярной формации играют гуморальные факторы,к которым она высоко чувствительна.
Ретикулярная формация состоит из двух частей: нисходящей и восходящей. По нисходящим путям (см. раздел средний и промежуточный мозг) ретикулярной формации, которые идут к спинному мозгу передаются импульсы либо облегчающие, активирующие, либо тормозящие функцию скелетных мышц. Нисходящая система влияет на функцию внутренних органов через нейроны вегетативной нервной системы (регуляция деятельности сердца, сосудов, органов пищеварения, дыхания, выделения и т.д.). Раздражение сетчатых образований среднего и промежуточного мозга усиливает выделение гормонов передней доли гипофиза, в том числе гонадотропных. При раздражении этих образований у крольчих и кошек течка не наступает.
Ретикулярной формации отводится важная роль в механизмах сна и бодрствования, а также в механизмах формирования целостных, и в частности, условнорефлекторных реакций организма.
По восходящим путям ретикулярной формации в кору больших полушарий передаются активирующие импульсы (неспецифическая афферентная система). Неспецифическая система действует на кору преимущественно диффузно: ее окончания разбросаны по всей коре.
Полная перерезка ретикулярной формации у животных вызывает кому и затем смерть. Влияя на кору больших полушарий, ретикулярная формация сама испытывает постоянные воздействия коры. Благодаря этому в кору не приходят ненужные импульсы, они блокируются на уровне ретикулярной формации.
Базальные, или подкорковые ядра (nuclei basales) - подкорковые образования, расположенные в белом веществе между корой больших полушарий и зрительными буграми. К ним относят хвостатое ядро, бледный шар, скорлупу, миндалину, безымянную субстанцию и другие, которые образуют стриопаллидарную систему. Она входит в состав экстрапирамидной системы и участвует в организации и построении движений. Миндалина является частью лимбической системы и вовлечена в формирование эмоций и памяти. Базальные ганглии участвуют в формировании инстинктов, которые служат врожденными формами поведения.
Физиологическую основу инстинктов составляют сложнейшие безусловноые цепные рефлексы, центры которых лежат в базальных ганглиях (полосатое тело) и ядрах промежуточного мозга.
Инстинкты - врожденные рефлексы, обеспечивающие поведение животных на различных стадиях их эволюционного и онтогенетического развития. Эти врожденные рефлексы обеспечивают на определенном этапе развития не только существование животных, но и их рост и дальнейшее развитие.
Инстинкты возникли на различных стадиях развития животного мира. Все инстинкты можно разделить на следующие основные группы:
1. Пищевые инстинкты - в эту группу включаются все рефлексы при помощи которых обеспечивается поиск и использование пищи. Каковы же главные причины, заставляющие животное искать пищу? В первую очередь это аппетит и чувство голода, которые возникают в результате голодных движений желудка и воздействия “голодной” крови на пищевые центры.
Главным, пусковым механизмом для пищеварительных инстинктов, для запука целой цепи рефлекторных актов является ощущение голода. Как только пищевой рефлекс прекращается, это порождает новую цепь рефлексов, связанных с обменом веществ.
Пищевые инстинкты включают целую цепь рефлекторных актов: разыскивания и преследование жертвы хищником, нападения, насыщения, переваривания и усвоения пищи.
2. Половые и родительские инстинкты - это врожденные рефлексы, которые обеспечивают размножение вида и сохранение потомства (инстинкт насиживания, сосания и др.).
3. Ориентировочные инстинкты - обеспечивают ориентировку животных в окружающей среде (поворот головы, настораживание ушей на малейший шорох, ориентировка в пространстве и др.). Эти инстинкты играют важную роль в сохранении жизни животного.
4. Комфортные инстинкты связаны с очисткой поверхности тела, движением отряхивания, чесания, движениями при купании (в воде, песке, золе). Они связаны с видом животных.
5. Оборонительные инстинкты - это комплекс безусловных рефлексов, которые защищают организм от раздражителя и воздействия факторов среды. Среди них одно из главных мест занимают двигательные и оборонительные реакции.
6. Инстинкт борьбы с ограничениями, или инстинкт свободы проявляется у животных, попавших в необычно сложную для них ситуацию. Например, дикие животные пытаются освободится от привязи зубами и лапами. Иногда этот инстинкт настолько сильно проявляется, что сильнейший пищевой рефлекс не тормозит его и животное (дикое) погибает в неволе не притронувшись к пище. У разных видов диких животных этот инстинкт проявляется неодинаково.
В нормальных условиях инстинкты представляют собой взаимодействие корковых и подкорковых центров: на сложные безусловнорефлекторные реакции наслаивается целый комплекс индивидуально приобретенных корковых реакций - условных рефлексов.








