 2015-06-14
2015-06-14 8333
8333Торможение может развиваться не только в постсинаптической мембране, но и в пресинаптическом отделе путем угнетения процесса высвобождения медиатора возбуждающими нервными окончаниями. В этом случае свойства постсинаптической мембраны не изменяются. Пресинаптическое торможение обнаружено в различных отделах ЦНС, но наиболее  часто оно встречается в мозговом стволе и особенно в спинном мозге.
часто оно встречается в мозговом стволе и особенно в спинном мозге.
Пресинаптическое торможение возникает в пресинаптической области перед синаптическим контактом, т.е. там, где аксон возбуждающего нейрона не имеет миелиновой оболочки перед переходом его в нервное окончание (рис. 78). С этим лишенным миелина участком контактирует аксон тормозного вставочного нейрона и образует тормозной аксоаксональный синапс. Импульсы, поступающие по аксону тормозного нейрона в аксоаксональный синапс, вызывают выделение тормозного медиатора (вероятнее всего гамма-аминомасляной кислоты), который вызывает сильную деполяризацию окончания аксона возбуждающего нейрона. Деполяризованный участок аксона теряет способность проводить нервные импульсы к синапсу и, следовательно, ко второй нервной клетке.
Пресинаптическое торможение особенно эффективно при обработке информации, поступающей к нейрону по различным пресинаптическим путям. В этом случае возбуждение, поступающее по одному из синаптических входов, может быть избирательно уменьшено или даже полностью прекращено при отсутствии влияния на другие входы к нейрону. Сам постсинаптический нейрон остается незаторможенным и способен реагировать на импульсы, поступающие по другим входам. Пресинаптическое торможение обусловливает ограничение притока афферентных импульсов к нервным центрам.
Торможение деятельности нейрона может осуществляться и без участия особых тормозных структур. В этом случае торможение развивается в возбуждающих синапсах в результате сильной деполяризации мембраны под влиянием слишком частого поступления нервных импульсов в пресинаптический отдел. Такой тип торможения был изучен русским физиологом Н.Е. Введенским и назван пессимальным.
В основе развития пессимального торможения лежит стойкая деполяризация, которая может возникнуть как в постсинаптической, так и в пресинаптической мембране синапса под влиянием очень частого поступления сильных импульсов. К пессимальному торможению особенно склонны вставочные нейроны спинного мозга, нейроны ретикулярной формации и некоторые другие нервные клетки, в которых при частом ритмическом раздражении деполяризация синаптических мембран может быть сильной и стойкой.
5.12. Спинной мозг
Спинной мозг у человека является низшим отделом ЦНС, имеющим сегментарную организацию. Сегментарность заключается в том, что: 1) сегменты спинного мозга иннервируют участки тела, расположенные на том же уровне (шейные – шею, грудные – грудь и т.д.); 2) нейроны спинного мозга напрямую связаны с рабочими органами в отличие от нейронов надсегментарных центров, например коры больших полушарий, которые управляют деятельностью мышц через сегментарные центры, в том числе и через спинной мозг. Спинной мозг у человека имеет значительно меньшую самостоятельность по сравнению со спинным мозгом животных. У человека его вес по отношению к головному мозгу составляет всего 2 % (у кошек – 25 %, у кролика – 45 %).
Спинной мозг выполняет интегративную, проводниковую и рефлекторную функции. Интегративная функция заключается в объединении (интеграции) поступающей информации от рецепторов. Она у спинного мозга проявляется слабо. Интегративная функция более значительна у вышележащих отделов ЦНС и наиболее выражена у коры больших полушарий.
Проводниковая функция у спинного мозга выражена очень хорошо и осуществляется благодаря проводящим путям, расположенным в белом веществе. Различают восходящие и нисходящие проводящие пути. По восходящим путям импульсы поступают от нижележащих сегментов спинного мозга к вышележащим сегментам и к головному мозгу, а по нисходящим путям – в обратном направлении.
Рефлекторная функция спинного мозга осуществляется благодаря передаче нервных импульсов с афферентных путей на эфферентные, в результате чего происходит ответная реакция на раздражение. Связь спинного мозга с рецепторами и рабочими органами осуществляется посредством нервных волокон, входящих в спинной мозг и выходящих из него в составе спинномозговых корешков. Распределение функций этих волокон подчиняется закону Белла-Мажанди, а именно: все чувствительные (афферентные) волокна входят в спинной мозг через задние корешки, а все двигательные и вегетативные (эфферентные) волокна выходят через передние корешки. В задних корешках волокон примерно в пять раз больше, чем в передних. По задним корешкам в спинной мозг поступают импульсы от рецепторов скелетных мышц, сухожилий, кожи, сосудов, внутренних органов. Передние корешки содержат волокна к скелетным мышцам и вегетативным ганглиям. В вегетативных ганглиях происходит синаптическая передача импульсов на нейроны, аксоны которых иннервируют внутренние органы.
Спинной мозг выполняет много рефлекторных функций. В нем находятся центры, осуществляющие движения всех скелетных мышц за исключением мышц, расположенных на голове. Также в спинном мозге расположены центры всех рефлексов мочеполовой системы и прямой кишки, терморегуляторных рефлексов, центры, регулирующие обмен веществ, сокращение диафрагмы, центры большинства сосудистых рефлексов. Кроме того, спинной мозг постоянно посылает нервные импульсы к мышцам, поддерживая их небольшое напряжение и готовность к работе – мышечный тонус.
Степень проявления рефлексов зависит от наличия связи структур спинного мозга со структурами головного мозга. После спинализации, т.е. отделения спинного мозга от головного у животных посредством перерезки или вследствие травмы у человека, исчезают многие сложные формы активности, создаваемые спинным мозгом. Такое состояние получило название спинальный шок. Продолжительность спинального шока у человека от нескольких недель до 4–5 месяцев. Когда шок проходит, рефлексы восстанавливаются.
В составе серого вещества спинного мозга человека насчитывают около 13,5 млн. нервных клеток. Из них мотонейронысоставляют всего 3 %, а 97 % являются вставочными нейронами. Рефлексы, протекающие с участием спинного мозга, подразделяются на двигательные, осуществляющиеся с помощью альфа-мотонейронов передних рогов, и вегетативные, осуществляемые эфферентными клетками боковых рогов и бокового промежуточного вещества.
Альфа-мотонейроны – это крупные клетки, от которых отходят толстые быстропроводящие нервные волокна. Они осуществляют передачу к скелетным мышцам импульсов, выработанных в спинном мозге. Аксон каждого мотонейрона, вступая в мышцу, многократно делится, и может охватывать своими окончаниями до нескольких сотен экстрафузальных мышечных волокон, образуя с ними двигательную, или нейромоторную единицу.
Импульсы, поступающие от альфа-мотонейронов, вызывают сокращение скелетных мышц. Потенциалы действия в альфа-мотонейроне возникают с частотой, не превышающей 200–300 имп/с, и со следовой гиперполяризацией длительностью от 50 до 150 мс. В зависимости от частоты воспроизведения импульсов и длительности следовой гиперполяризации альфа-мотонейроны разделяют на фазические (фазные) и тонические. Фазическими мотонейронами иннервируются более быстрые, «белые» мышцы, тоническими – более медленные «красные».
Второй тип мотонейронов спинного мозга – гамма-мотонейроны посылают тонкие волокна к интрафузальным (внутриверетенным) мышечным волокнам, которые являются мышечными рецепторами. Импульсы, поступающие от гамма-мотонейронов, не вызывают сокращение скелетных мышц, а изменяют чувствительность мышечных рецепторов.
В спинном мозге замыкается огромное количество рефлекторных дуг, с помощью которых регулируются функции организма. Типичные спинномозговые (спинальные) рефлексы, т.е. рефлексы, присущие самому спинному мозгу, можно изучить в чистом виде только после отделения спинного мозга от головного у животного (спинальное животное) или у человека, у которого в результате травмы произошёл разрыв спинного мозга на границе с продолговатым мозгом. Первым следствием нарушения целостности спинного мозга является спинальный шок. Он проявляется в резком падении возбудимости и в угнетении рефлекторных функций всех рефлекторных центров, расположенных ниже места перерезки или разрыва.
Спустя несколько недель, а то и месяцев у человека спинальный шок медленно проходит и постепенно восстанавливается рефлекторная деятельность скелетных мышц, величина кровяного давления, рефлексы мочеиспускания, дефекации и ряд половых рефлексов. Это свидетельствует о том, что центры этих рефлексов располагаются в спинном мозге.
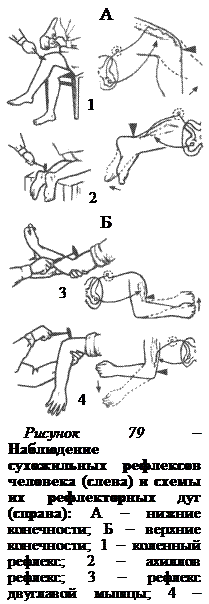
Первыми после отделения головного мозга от спинного и снятия шока появляются сгибательные и разгибательные рефлексы. Вслед за ними обнаруживаются сухожильные рефлексы (рис. 79), дуги которых являются моносинаптическими, т.е. имеющими только один синапс. Моносинаптическая дуга состоит из чувствительного нейрона, дендриты которого идут от рецепторов растяжения – мышечных веретён, и эфферентного нейрона, оканчивающегося на мышечных волокнах. В спинном мозге меньше моносинаптических дуг и больше полисинаптических.
Сухожильные рефлексы легко вызываются с помощью короткого удара по сухожилию и имеют важное диагностическое значение в неврологической практике. Ответная рефлекторная реакция на удар по сухожилию проявляется в виде резкого сокращения мышцы. Особенно выражены сухожильные рефлексы в мышцах разгибателях ноги (рис. 79, А), таких как четырехглавая мышца бедра (коленный рефлекс) или трехглавая мышца голени (ахиллов рефлекс). Сухожильные рефлексы можно вызвать и в мышцах-сгибателях. На руке они четко проявляются на двуглавой и трехглавой мышцах (рис. 79, Б).
 Важное значение для поддержания мышечного тонуса, выпрямленной позы и стояния человека имеют рефлексы растяжения. Под влиянием силы тяжести раздражаются рецепторы растяжения (мышечные веретёна) в мышце. Так, при стоянии четырехглавая мышца бедра подвергается растяжению из-за тенденции колена сгибаться под влиянием гравитации. От мышечных веретён четырехглавой мышцы поступают длительное время асинхронные импульсы к альфа-мотонейронам спинного мозга. В ответ альфа-мотонейроны посылают тоже асинхронные импульсы к экстрафузальным волокнам четырехглавой мышцы. В результате этого мышца отвечает плавным длительным сокращением, автоматически противодействующим силе тяжести. Рефлексы растяжения иногда относят к группе тонических или позных рефлексов, т.к. их объединяет длительное поддержание рефлекторного сокращения разгибательных и реже сгибательных мышц без заметного их утомления. Тонические рефлексы направлены на поддержание определенной позы. В их осуществлении кроме механизмов спинного мозга участвуют двигательные центры головного мозга.
Важное значение для поддержания мышечного тонуса, выпрямленной позы и стояния человека имеют рефлексы растяжения. Под влиянием силы тяжести раздражаются рецепторы растяжения (мышечные веретёна) в мышце. Так, при стоянии четырехглавая мышца бедра подвергается растяжению из-за тенденции колена сгибаться под влиянием гравитации. От мышечных веретён четырехглавой мышцы поступают длительное время асинхронные импульсы к альфа-мотонейронам спинного мозга. В ответ альфа-мотонейроны посылают тоже асинхронные импульсы к экстрафузальным волокнам четырехглавой мышцы. В результате этого мышца отвечает плавным длительным сокращением, автоматически противодействующим силе тяжести. Рефлексы растяжения иногда относят к группе тонических или позных рефлексов, т.к. их объединяет длительное поддержание рефлекторного сокращения разгибательных и реже сгибательных мышц без заметного их утомления. Тонические рефлексы направлены на поддержание определенной позы. В их осуществлении кроме механизмов спинного мозга участвуют двигательные центры головного мозга.
Ритмические рефлексы, или рефлексы мышц-антагонистов, лежат в основе правильных попеременных движений при ходьбе, беге, чесании и т.д. Эти рефлексы характеризуются тем, что при возбуждении мотонейронов мышц-сгибателей одновременно происходит торможение мотонейронов мышц-разгибателей. При этом в конечности другой стороны наблюдаются обратные явления. Центры рефлекса ходьбы лежат в поясничных сегментах спинного мозга.
К двигательным рефлексам спинного мозга относят также висцеромоторные рефлексы. Они возникают при определенных раздражениях внутренних органов и характеризуются появлением двигательных реакций мышц грудной и брюшной стенки, мышц-разгибателей спины (например, при икоте).
Двигательные рефлексы спинного мозга называют также соматическими, т.к. они проявляются в деятельности скелетных мышц. Кроме их осуществления, спинной мозг играет важную роль в рефлекторной регуляции внутренних органов, являясь центром многих вегетативных рефлексов. Вегетативные рефлексы спинного мозга разделяют на симпатические и парасимпатические. В боковых рогах сегментов грудного отдела и верхних сегментов поясничного отдела спинного мозга расположены центры симпатического отдела, а в боковом промежуточном веществе крестцового отдела – центры парасимпатического отдела вегетативной нервной системы. Эти центры участвуют в регуляции пищеварения, дыхания, кровообращения, мочеиспускания, дефекации, осуществляют некоторые половые рефлексы.
5.13. Функции продолговатого мозга и варолиева моста
Продолговатый мозг является продолжением спинного и в нижней части похож на него по строению и форме. В продолговатом мозге частично сохраняется сегментарное строение, типичное для спинного мозга.
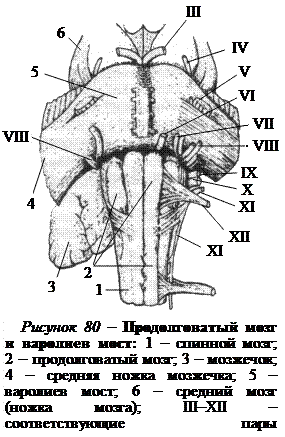
Из 12 пар черепных (черепно-мозговых) нервов, выходящих из головного мозга и черепа самостоятельно, без связи со спинным мозгом, V–XII пары отходят от ядер, расположенных в продолговатом мозге и мосту (рис. 80). Часть из этих нервов иннервируют лицевую и глазодвигательную мускулатуру, что свидетельствует о сегментарных чертах строения продолговатого мозга и моста.
Продолговатый мозг и мост выполняют проводниковую, рефлекторную и интегративную функции. В них поступают афферентные нервные волокна от вестибулярных и слуховых рецепторов, от кожи и мышц головы, от внутренних органов. Проводниковая функция продолговатого мозга и варолиева моста заключается в передаче импульсов из спинного мозга в вышележащие отделы ЦНС и в проведении импульсов, идущих из головного в спинной мозг. В продолговатом мозге заканчивается кортико-бульбарный нисходящий путь, по которому импульсы от коры больших полушарий поступают к ядрам черепно-мозговых нервов. От продолговатого мозга к спинному отходят нисходящие ретикуло-спинальные пути и вестибуло-спинальный пучок.
 Через продолговатый мозг проходят восходящие пути от рецепторов слуховой и вестибулярной чувствительности. В продолговатом мозге афферентные нервные волокна, несущие информацию от рецепторов кожи и мышечных рецепторов, образуют синапсы на вторых нейронах, аксоны которых составляют путь в таламус.
Через продолговатый мозг проходят восходящие пути от рецепторов слуховой и вестибулярной чувствительности. В продолговатом мозге афферентные нервные волокна, несущие информацию от рецепторов кожи и мышечных рецепторов, образуют синапсы на вторых нейронах, аксоны которых составляют путь в таламус.
Продолговатый мозг и мост выполняют сложные рефлекторные функции, в том числе и жизненно важные для организма. На дне IV-го желудочка в ретикулярной формации продолговатого мозга находятся жизненно важные дыхательный центр, регулирующий дыхание, и сосудодвигательный центр, регулирующий работу сердца и просвет кровеносных сосудов. В продолговатом мозге и мосту находится ряд рефлекторных центров, связанных с процессами пищеварения, а именно: центры жевания, сосания, слюноотделения, глотания, выделения пищеварительных соков желудком, поджелудочной железой и др. Здесь же находятся центры некоторых защитных рефлексов: чихания, кашля, рвоты, мигания, слезоотделения.
Целая группа рефлексов продолговатого мозга и моста направлена на поддержание позы тела. В зависимости от рецепторов, при возбуждении которых осуществляется поддержание позы, эти рефлексы подразделяют на шейные тонические рефлексы и вестибулярные (лабиринтные). Шейные тонические рефлексы запускаются при возбуждении проприорецепторов мышц шеи вследствие изменения положения головы относительно туловища.
Вестибулярные рефлексы неразрывно связаны с шейными тоническими рефлексами и дополняют их, т.к. не зависят от положения головы относительно туловища. Вестибулярные рефлексы подразделяются на статические и статокинетические. Статические рефлексы вызываются главным образом возбуждением рецепторов, расположенных в мешочке и маточке вестибулярного аппарата, и обеспечивают поддержание позы и равновесия тела при разнообразных неподвижных его положениях в пространстве. Среди статических рефлексов различают две группы: 1) позно-тонические рефлексы, или рефлексы положения, благодаря которым сохраняется определенная поза; 2) установочные, или выпрямительные рефлексы, которые обеспечивают переход из неестественной позы в обычное положение.
Статокинетические рефлексы обеспечивают поддержание позы тела при изменении скорости движения. Эти рефлексы вызываются при возбуждении рецепторов, расположенных в полукружных каналах вестибулярного аппарата. Возбуждение же рецепторов возникает при движении эндолимфы в каком-либо из трех полукружных каналов в результате ускорения или торможения тела человека. К статокинетическим вестибулярным рефлексам относятся также лифтные рефлексы, которые проявляются в увеличении тонуса мышц сгибателей и уменьшении тонуса мышц разгибателей при ускорении вверх. При ускорении вниз наблюдается обратная картина: тонус мышц сгибателей уменьшается, а тонус мышц разгибателей повышается. В результате в начале подъема происходит сгибание конечностей и человек слегка приседает, а в начале спуска – их разгибание и человек слегка выпрямляется.
5.14. Средний мозг
Средний мозг совместно с варолиевым мостом и продолговатым мозгом образует ствол головного мозга, в котором он занимает относительно небольшую по объему часть. К стволу мозга иногда также относят промежуточный мозг и мозжечок. Средний мозг расположен над варолиевым мостом спереди от мозжечка. Он состоит из ножек мозга и крыши среднего мозга, или четверохолмия (рис. 81).
Средний мозг выполняет рефлекторную, интегративную и проводниковую функции. Он участвует в осуществлении целого ряда важных безусловных рефлексов. Так, при участии бугров четверохолмия осуществляются ориентировочные зрительные и слуховые рефлексы на неожиданные раздражители. В ответ на внезапное раздражение происходит поворот головы и глаз в сторону раздражителя, а у животных – настораживания ушей. Этот рефлекс И.П. Павлов назвал рефлексом «Что такое?». Он необходим для подготовки организма к своевременной реакции на любое новое воздействие. Он сопровождается усилением тонуса мыщц-сгибателей (подготовка к двигательной реакции) и изменениями вегетативных функций (дыхание, сердцебиения).
 Передние бугры четверохолмия являются первичными подкорковыми зрительными центрами, а задние бугры – первичными подкорковыми слуховыми центрами. Ядра, расположенные в передних и задних буграх четверохолмия, участвуют в осуществлении сторожевого рефлекса при внезапной подаче светового или звукового раздражителей. Этот рефлекс проявляется в перераспределении мышечного тонуса: происходит повышение тонуса мышц-сгибателей и уменьшение тонуса мышц-разгибателей, что мобилизует организм на быструю ответную реакцию.
Передние бугры четверохолмия являются первичными подкорковыми зрительными центрами, а задние бугры – первичными подкорковыми слуховыми центрами. Ядра, расположенные в передних и задних буграх четверохолмия, участвуют в осуществлении сторожевого рефлекса при внезапной подаче светового или звукового раздражителей. Этот рефлекс проявляется в перераспределении мышечного тонуса: происходит повышение тонуса мышц-сгибателей и уменьшение тонуса мышц-разгибателей, что мобилизует организм на быструю ответную реакцию.
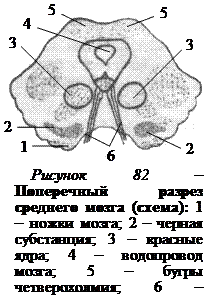 На поперечном разрезе среднего мозга видно, что каждая ножка состоит из покрышки и основания, на границе которых находится черное вещество, или черная субстанция (рис. 82). Черная субстанция представляет собой скопление тел нейронов, содержащих пигмент меланин, придающий нейронам характерный темный цвет.
На поперечном разрезе среднего мозга видно, что каждая ножка состоит из покрышки и основания, на границе которых находится черное вещество, или черная субстанция (рис. 82). Черная субстанция представляет собой скопление тел нейронов, содержащих пигмент меланин, придающий нейронам характерный темный цвет.
Черная субстанция среднего мозга участвует в регуляции тонуса мышц и различных тонких движений (например, мелких движений пальцев рук). Черная субстанция функционально связана с подкорковыми ядрами больших полушарий – полосатым телом и бледным шаром. Нейроны черной субстанции имеют дофаминергическую природу, т.к. синтезируют медиатор дофамин. Аксоны этих нейронов идут к полосатому телу и образуют синапсы на его нервных клетках, также содержащих в значительных количествах дофамин. Повреждение черной субстанции приводит к дегенерации дофаминергических нервных путей к полосатому телу, что является одной из причин возникновения тяжелого заболевания нервной системы – болезни Паркинсона.
В покрышке среднего мозга располагаются ядра глазодвигательного и блокового нервов (соответственно III-й и IV-й пары черепномозговых нервов). Эти нервы управляют движениями глаз, аккомодацией глаза, зрачковым рефлексом.
Важным промежуточным центром проводящих путей ствола мозга являются красные ядра (рис. 82). Они связаны нервными путями с корой больших полушарий, ретикулярной формацией, мозжечком и спинным мозгом. От красных ядер берет начало нисходящий руброспинальный путь, идущий к мотонейронам спинного мозга. С его помощью осуществляется регуляция тонуса скелетных мышц, а именно: происходит повышение тонуса нейронов, иннервирующих мышцы-сгибатели, и уменьшение тонуса нейронов мышц-разгибателей. Это имеет большое значение при поддержании позы и осуществлении движений. Если у животного перерезать головной мозг на уровне задних бугров четверохолмия так, чтобы красные ядра оказались выше линии перерезки, то у такого животного перестают поступать импульсы от красных ядер к нейронам мышц-сгибателей и наступает резкое повышение тонуса разгибательной мускулатуры. Конечности у животного сильно выпрямляются, голова запрокинута назад, хвост приподнят, конечности трудно согнуть даже силой. Животное не может принять нормальную позу. Такое состояние мускулатуры тела называется децеребрационной ригидностью (рис. 83). Если поставить такое животное на лапы, то оно сохраняет стоячее положение, так как тонус разгибателей настолько высок, что сгибания в суставах не происходит.
 Предполагается, что красные ядра являются не только источником, но и посредником в регуляции разгибательных рефлексов, передавая тормозные влияния от мозжечка и моторных областей коры. Удаление мозжечка или моторных областей коры приводит к усилению тонуса мышц-разгибателей.
Предполагается, что красные ядра являются не только источником, но и посредником в регуляции разгибательных рефлексов, передавая тормозные влияния от мозжечка и моторных областей коры. Удаление мозжечка или моторных областей коры приводит к усилению тонуса мышц-разгибателей.

