 2015-06-16
2015-06-16 4841
4841Для нейтрализации токсических форм O2 существующие прокариоты выработали различные защитные механизмы, которые могут быть разделены на несколько типов. В основе систем защиты первого типа лежит активность специальных ферментов, для которых разложение токсических форм O2 является основной и в ряде случаев единственной функцией. В системах защиты второго типа для разрушения токсических форм O2 используются определенные клеточные метаболиты. Как правило, в этом случае участие в защите клетки от токсических эффектов производных O2 является не единственной функцией этих метаболитов. Наконец, к защитным механизмам особого типа может быть отнесен ряд приспособлений, выработанных прокариотами на разных уровнях: популяционном, физиологическом, структурном. Более вероятно, что они были созданы для других целей, но оказались полезными и для детоксикации O2.
Ферментные системы защиты. Передовой линией защиты от токсического действия производных O2 являются ферменты: супероксиддисмутаза, захватывающая молекулы О2 (реакция 4), каталаза и пероксидаза, улавливающие H2O2:
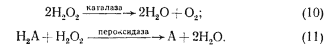
Они сводят до минимума концентрацию в клетке O2–. и H2O2 и не дают им возможности взаимодействовать с образованием ОН. (реакция 5).
Супероксиддисмутаза обнаружена у хемотрофных прокариот, использующих O2 (облигатно и факультативно аэробных форм), а также у изученных представителей из групп фотосинтезирующих прокариот. Среди анаэробов фермент найден у подавляющего большинства аэротолерантных форм. Исключение составляют некоторые молочнокислые бактерии, однако в клетках большинства из них содержатся высокие концентрации (до 30 мМ) ионов двухвалентного марганца. Оказалось, что Мп2+, для которого показана способность окисляться под действием O2, в таких концентрациях способен так же эффективно убирать образующиеся супероксидные ионы, как это делает супероксиддисмутаза, содержание которой в клетке обычно поддерживается на микромолярном уровне. Таким образом, у этих молочнокислых бактерий функцию нейтрализации O2 выполняют ионы Mn2+.
В клетках некоторых видов молочнокислых бактерий не найдено ни супероксиддисмутазы, ни высоких концентраций Mn2+. Эти виды характеризуются очень высокой чувствительностью к O2.
Среди облигатных анаэробов супероксиддисмутаза обнаружена у многих представителей рода Clostridium. Изучение их устойчивости к O2 обнаруживает четкую связь с содержанием в клетках этого фермента. Виды, имеющие супероксиддисмутазу, характеризуются умеренной или даже высокой устойчивостью к O2 по сравнению с видами, у которых этот фермент отсутствует. Супероксиддисмутаза найдена у разных видов строго анаэробных бактерий. Число организмов с не выявленной до сих пор супероксиддисмутазой очень мало.
Обнаружение супероксиддисмутазы у строгих анаэробов (гораздо более распространенное, чем предполагали раньше) ставит вопрос о ее физиологической роли у этих организмов. Способность последних расти только в бескислородной среде делает неясным функции фермента в данных условиях. Возможно, что только при попадании строгого анаэроба в неблагоприятные для него аэробные условия синтез фермента индуцируется молекулярным кислородом, что и обеспечивает организму защиту от O2 в этих условиях.
Супероксиддисмутаза — фермент, содержащий в активном центре в качестве простетической группы ионы металла. У прокариот — это атомы марганца и/или железа. Большинство изученных супероксиддисмутаз построено из двух идентичных субъединиц, каждая из которых содержит по одному атому металла. Fe- и Mn-ферменты сходны по аминокислотной последовательности. Попытки выявить связь между физиологическими и иными особенностями организмов и металлоформой содержащегося в них фермента не привели к определенному заключению. И та и другая формы супероксиддисмутазы обнаружены у представителей грамположительных и грамотрицательных прокариот, среди фото- и хемотрофов, облигатных анаэробов, аэробов и факультативно анаэробных форм. Более того, обе металлоформы супероксиддисмутазы могут присутствовать у одного организма и даже входить в состав молекулы одного фермента. Для некоторых видов показано, что синтез того или иного типа фермента зависит от наличия ионов металла в среде культивирования.
Супероксиддисмутаза изученных хемотрофных прокариот — не связанный с мембранами фермент, локализованный в цитоплазме. У Е. coli, в клетках которой обнаружены Fe-, Mn- и Fe/Mn-формы фермента, Fe-супероксиддисмутаза локализована в периплазматическом пространстве, а Mn-содержащий фермент — в цитоплазме. В связи с этим высказывается предположение, что металлоформы фермента играют разную роль в защите клетки от O2–.: Fe-содержащий фермент защищает клетку от экзогенных супероксидных анионов, а Mn-содержащий — от эндогенных.
Особо остро стоит проблема защиты от молекулярного кислорода и его производных в клетках цианобактерий. Вероятно, именно они впервые в наибольшей степени ощутили последствия токсических эффектов кислорода. Супероксиддисмутаза найдена у всех цианобактерий. В клетках Anacystis nidulans (Synechococcus) Fe-супероксиддисмутаза, составляющая до 90% от общего количества фермента, локализована в цитозоле клетки, а Mn-содержащая форма — в тилакоидах. Функция последней формы фермента сводится, вероятно, к перехвату ионов O2–., возникающих в процессе фотосинтетического электронного транспорта.
Каталаза и пероксидаза. Перекись водорода разрушается двумя классами родственных ферментов, катализирующих ее двухэлектронное восстановление до H2O и использующих в качестве донора электронов H2O2 в случае каталазы (реакция 10) или различные органические соединения в случае пероксидазы (реакция 11).
Каталазная и пероксидазная активности обнаружены у всех облигатно и факультативно аэробных прокариот. Среди облигатных анаэробов эти ферменты распространены значительно в меньшей степени, чем супероксиддисмутаза. Обнаружены многие строгие и аэротолерантные анаэробы, содержащие супероксиддисмутазу, но не содержащие каталазы. К их числу можно отнести и те молочнокислые бактерии, у которых дисмутация образующихся ионов O2–. обеспечивается Mn2+, находящимся в клетках в высоких концентрациях.
Отсутствие каталазы у молочнокислых бактерий связано с тем, что они не могут синтезировать гем — простетическую группу фермента, но способны к синтезу апофермента. При добавлении гемовых групп извне молочнокислые бактерии образуют гемсодержащую каталазу. У ряда молочнокислых бактерий обнаружена каталаза, не содержащая гемовой группы, названная поэтому псевдокаталазой. Выделенный фермент состоит из шести идентичных полипептидных цепей, соединенных между собой нековалентными силами. Каждая субъединица содержит 1 атом марганца.
Перекись водорода, возникающая в результате взаимодействия клеток с O2, устраняется и неферментативными путями. Известно, что ионы Fe2+ в водном растворе ускоряют восстановление H2O2 до H2O. В клетке всегда содержится некоторое количество ионов железа. Разрушение H2O2 может происходить и за счет выделяющихся в культуральную среду восстановленных веществ.
Для анаэробных прокариот, способных переносить контакт с O2 и его производными в относительно небольших масштабах, необходимо присутствие в клетках супероксиддисмутазы, "убирающей" O2–.. Наличие каталазы при этом не обязательно, поскольку возникающая в реакции дисмутации и других реакциях перекись водорода разлагается спонтанно или с участием неферментативных катализаторов, и организмы в целом справляются с ней в этих условиях. Таким образом, при осуществлении энергетического метаболизма анаэробного типа для устранения токсических эффектов O2. достаточно одной ферментной преграды в виде супероксиддисмутазы.
Резкое возрастание масштабов взаимодействия прокариот с O2 при функционировании метаболизма аэробного типа делает неэффективными неферментативные пути устранения H2O2. Для разложения перекиси водорода, образующейся в больших количествах, необходимы специальные ферменты, повышающие скорость разложения H2O2 на несколько порядков. Это обеспечивается каталазой и пероксидазой. Таким образом, в условиях активного взаимодействия клеток с O2, делающего возможным аэробную жизнь, система ферментной защиты от его токсических эффектов сформирована с участием супероксиддисмутазы, каталазы и пероксидазы в качестве необходимых компонентов (рис. 87).
Механизмы защиты с помощью клеточных метаболитов. Защита против одного из самых токсичных производных O2 — синглетного кислорода — осуществляется с помощью разных биологически важных молекул. Все виды тушения *O2 можно разделить на физические и химические. Физическим называют тушение, которое не приводит к разрушению тушителя:
*O2 + A O2 + A
Химическое тушение приводит к окислению тушителя:
*O2 + A O2 + Aок
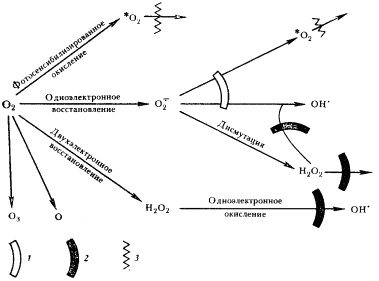 |
| Рис. 87. Системы клеточной защиты от токсических производных молекулярного кислорода: 1 — супероксиддисмутаза, ионы Mn2+; 2 — каталаза, пероксидаза; 3 — тушение каротиноидами |
Преимущественно по химическому механизму тушение *O2 осуществляется насыщенными жирными кислотами, липидами, аминокислотами, нуклеотидами и другими соединениями. Механизмы химического тушения разнообразны, но в большинстве случаев начальной стадией является образование лабильной циклической перекиси с последующим ее разложением, которое приводит к возникновению свободных радикалов. Химическое тушение *O2 может приводить в клетке к существенным деструктивным последствиям. К тушению в основном по физическому механизму способны молекулы разных химических соединений. Наиболее эффективны в этом отношении каротиноиды, широко распространенные в мире прокариот. Они обнаружены в клетках многих аэробных хемотрофов, являются обязательным компонентом пигментного аппарата всех фототрофов. В клетках фотосинтезирующих организмов, как отмечалось выше, активными фотосенсибилизаторами являются хлорофиллы. Однако возможность фотоокислительных эффектов в условиях функционирования фотосинтетического аппарата довольно низка, во-первых, из-за чрезвычайно короткого (10–11 с) времени пребывания хлорофилла в возбужденном состоянии и, во-вторых, из-за защиты клеток от фотоокисления каротиноидами.
Впервые роль каротиноидов в предотвращении летального эффекта, вызываемого фотоокислением, была показана при изучении бескаротиноидного мутанта пурпурной бактерии Rhodopseudomonas sphaeroides. Исходная культура хорошо росла фототрофно в анаэробных условиях, но могла также расти на свету, и в темноте в аэробных условиях. Полученный из нее мутант, лишенный каротиноидов, обладал низкой скоростью роста на свету в анаэробных условиях и в темноте в аэробных условиях, но быстро погибал при перенесении на свет + воздух.
Фотоокислительные повреждения могут развиваться и у нефотосинтезирующих прокариот, так как в их клетках также имеются окрашенные молекулы, поглощающие видимый свет, которые могут функционировать как фотосенсибилизаторы. Действие каротиноидов не ограничивается только их участием в защите от фотодинамического эффекта. Они гасят синглетное состояние кислорода независимо от того, в каких реакциях он возникает: на свету или в темноте.
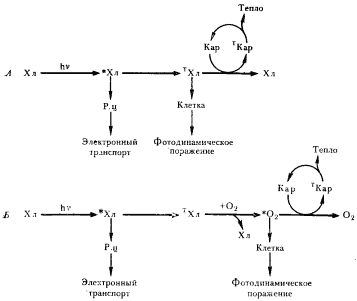 |
| Рис. 88. Механизмы защитного действия каротиноидов. А — тушение каротиноидами триплетного состояния хлорофилла. Б — тушение каротиноидами синглетного кислорода: *Хл — молекула хлорофилла в возбужденном синглетном состоянии; TХл — молекула хлорофилла в возбужденном триплетном состоянии; TKap — молекула каротиноида в возбужденном триплетном состоянии; Р. ц. — реакционный центр |
Механизм защитного действия каротиноидов у фотосинтезирующих организмов заключается в следующем (рис. 88). Молекула хлорофилла, поглотившая свет, быстро (10–12 с) переносит энергию синглетного возбужденного состояния в реакционный центр. Из 104 поглощенных квантов света приблизительно 4 приводят к переходу молекулы хлорофилла в возбужденное триплетное состояние. Возникает возможность фотодинамического поражения. Каротиноиды могут участвовать в трех защитных реакциях: 1) непосредственно тушить триплетное состояние хлорофилла, переводя его в основное состояние (рис. 88, А); возникающая при этом триплетная молекула каротиноида отдает избыточную энергию в виде тепла и возвращается в основное состояние; 2) триплетный хлорофилл не гасится каротиноидами; происходит его взаимодействие с O2, переводящее последний в возбужденное синглетное состояние; синглетный кислород гасится каротиноидами (рис. 88, Б); 3) синглетный кислород, не подвергшийся гашению каротиноидами по физическому механизму, может взаимодействовать с ними в химической реакции, приводящей к окислению каротиноидов. Участие каротиноидов в любой из трех описанных выше реакций будет снижать уровень образования в клетке *O2.
Приспособления прокариот, помогающие им в защите от токсических эффектов молекулярного кислорода. В клетках тех облигатно анаэробных клостридиев, у которых не обнаружено ни супероксиддисмутазы, ни каталазы, доступным средством нейтрализации O2 служит вытеснение его из среды культивирования с помощью активно выделяющихся газообразных продуктов (CO2 и H2), сопровождающих брожение, а также поглощение клеточной суспензией кислорода из среды, приводящее к гибели части клеток, но дающее возможность оставшимся размножаться в условиях пониженного содержания O2.
Молочнокислыми бактериями в направлении защиты от молекулярного кислорода сделан определенный шаг вперед. Эти бактерии — единственная группа прокариот, не имеющих гемсодержащей каталазы, способных расти в присутствии воздуха. Поиски механизмов нейтрализации O2 и его производных привели к обнаружению у них помимо супероксиддисмутазы и высокой внутриклеточной концентрации ионов Mn2+, осуществляющих разложение O2–., псевдокаталазы, а также каталазо- и пероксидазоподобной активности. У отдельных представителей молочнокислых бактерий просматривается более четко выраженная степень приспособленности к O2, приводящая к попыткам определенного его полезного использования. Для некоторых молочнокислых бактерий рода Lactobacillus показано ускорение гликолитического разложения глюкозы в аэробных условиях. Это связано с тем, что в аэробных условиях водород с НАД-H2 может прямо передаваться на O2, освобождая часть пировиноградной кислоты от ее акцепторной функции, как это происходит при обычном молочнокислом брожении. Освобожденная от этой "обязанности" пировиноградная кислота может теперь окисляться до ацетил-КоА, последующее метаболизирование которого до ацетата приводит к синтезу молекулы АТФ. Как можно видеть, участие кислорода в этом процессе прямо не связано с получением клеткой энергии (при передаче водорода с НАД-H2 на O2 энергия в форме АТФ не образуется), т. е. вся энергия получается за счет субстратного фосфорилирования, но O2, беря на себя акцепторную функцию, освобождает часть пирувата, которая может использоваться клеткой по энергетическому пути, что в конечном итоге приводит к повышению энергетического выхода брожения. Таким образом, прямое окисление части восстановленных переносчиков электронов в процессе брожения может иметь не только отрицательные, но и положительные последствия.
Как отмечалось выше, очень чувствителен к O2 процесс азотфиксации. Несмотря на это, способность фиксировать N2 широко распространена среди прокариот, различающихся отношением к молекулярному кислороду; она присуща хемотрофам и фототрофам, в том числе цианобактериям, осуществляющим кислородный фотосинтез. Фиксировать N2 могут свободноживущие формы и прокариоты, находящиеся в симбиозе с эукариотными организмами.
Изучение средств защиты этого процесса у прокариот показало, что в большинстве случаев она далека от 100%-ной эффективности. Среди аэробных азотфиксаторов можно выделить лишь немногие организмы, способные расти в среде с N2 в условиях равновесия с воздухом. Большинство может расти и фиксировать N2 только в условиях пониженной концентрации молекулярного кислорода, т. е. в микроаэробных условиях. Защита нитрогеназы в клетках факультативных анаэробов еще менее эффективна: они могут осуществлять активно фиксацию азота только в анаэробных условиях.
К числу аэробных азотфиксаторов относятся представители рода Azotobacter, у которых обнаружены различные защитные приспособления. Одно из них связано с резким увеличением дыхательной активности клеток, осуществляющих азотфиксацию в аэробных условиях. Дыхание в значительной мере служит в этом случае для "связывания" внутриклеточного O2. При этом обнаружены существенные перестройки как в клеточном строении азотобактера, выражающиеся в интенсивном развитии системы внутрицитоплазматических мембран, так и в организации самой дыхательной цепи, локализованной в этих мембранах. Дыхательная цепь Azotobacter vinelandii достаточно сложная, имеет разветвления на путях переноса электронов на уровне цитохрома b. Транспорт электронов, сопряженный с фосфорилированием, происходит по пути:
цитохромы b c4 c5 a1.
При осуществлении "дыхательной" защиты возрастает активность электронного транспорта по ветви: цитохромы b d, не связанной с запасанием энергии. Это приводит к тому, что несмотря на возрастание общей активности дыхания сопряжение электронного транспорта с запасанием энергии снижается. Таким образом, происходит "сжигание" части углеродных субстратов, которые используются для восстановления O2, без запасания при этом клеткой энергии.
В дополнение к вынужденному "принесению в жертву" части источников углерода высокие концентрации O2 вызывают в клетке обратимые изменения структуры нитрогеназы, делающие чувствительные к молекулярному кислороду участки менее доступными для него. Высказываются разные предположения относительно того, как осуществляется "конформационная" защита. Возможно, при этом происходит изменение взаимного расположения двух нитрогеназных белков. Не исключено участие в защите такого типа клеточной мембраны. Определенная стабилизация нитрогеназы в условиях высокой концентрации O2 происходит при добавлении к ферментному комплексу двухвалентных катионов. Наконец, обнаружены специальные защитные белки, образующие комплексы с нитрогеназными белками и приводящие к повышению их стабильности в присутствии O2. Никаких других функций, кроме защитной, у этих белков пока не найдено.
Большинство азотфиксирующих прокариот способны фиксировать молекулярный азот в микроаэробных условиях. К числу защитных приспособлений у них относятся: образование слизи, препятствующей диффузии в клетку O2 и тем самым создающей вокруг нее микроаэробную зону; формирование клеточных скоплений, затрудняющих доступ O2 к клеткам, расположенным внутри скопления, которым, таким образом, создаются более благоприятные условия для азотфиксации; существование азотфиксирующих видов в ассоциации с неазотфиксирующими аэробными гетеротрофами, защищающими нитрогеназу азотфиксаторов от доступа O2.
Специфические приспособления для защиты нитрогеназы от высоких концентраций O2 выработаны симбиотическими азотфиксаторами — клубеньковыми бактериями. Уже сами клубеньки, места активного размножения бактерий и фиксации ими N2, следует рассматривать как структуру, одним из назначений которой является ограничение доступа внутрь молекулярного кислорода. Эту же функцию выполняет содержащийся в клубеньках леггемоглобин (белок, аналогичный гемоглобину), способный активно связывать O2 и контролировать его поступление в бактероиды. В любом случае при осуществлении метаболизма аэробного типа дыхание также будет препятствовать накоплению в клетке молекулярного кислорода.
Наиболее остро стоит проблема защиты процесса азотфиксации от O2 в группе цианобактерий. У всех цианобактерий нитрогеназа чувствительна к O2, имеющему внеклеточное и внутриклеточное происхождение. В соответствии с этим у них можно выделить приспособления, направленные на защиту от экзогенного кислорода, и те, которые предназначены для нейтрализации O2. образующегося внутри клетки в процессе фотосинтеза.
Если все азотфиксирующие цианобактерий рассматривать под углом зрения степени защиты процесса азотфиксации от O2, то их можно разделить на две группы. К первой группе относятся цианобактерий, у которых защита азотфиксации от O2 наименее эффективна, поэтому вегетативные клетки могут фиксировать N2 только в анаэробных или микроаэробных условиях. Вторую группу составляют цианобактерий, у которых для осуществления азотфиксации в аэробных условиях сформированы специализированные клетки — гетероцисты.
У безгетероцистных цианобактерий защита нитрогеназы вегетативных клеток от O2, в первую очередь эндогенного, осуществляется с помощью разделения во времени процессов фотосинтеза и азотфиксации, непрерывного синтеза нитрогеназы, высокой активности супероксиддисмутазы в сочетании с каталазной и пероксидазной активностями. В центре филаментов некоторых безгетероцистных форм часто выделяются слабопигментированные вегетативные клетки, у которых предположительно подавлена способность к фотосинтетической фиксации CO2 и тем самым созданы более благоприятные условия для азотфиксации. (Это не гетероцисты, но, вероятно, именно из них впоследствии развились гетероцисты как центры азотфиксации в аэробных условиях). Средством защиты от экзогенного O2 служит синтез большого количества слизи, часто окружающей клетки азотфиксирующих цианобактерий. Существование в виде колониальных форм также может обеспечивать создание анаэробных условий для клеток, располагающихся в центральной части колонии.
Наиболее совершенна защита от эндогенного и экзогенного молекулярного кислорода в гетероцистах. Гетероцисты не способны к фотосинтетическому выделению O2. А высокие активности окислительного пентозофосфатного пути, поставляющего электроны в дыхательную цепь, где они акцептируются O2, повышенные уровни супероксиддисмутазы сравнительно с вегетативными клетками, образование гетероцистами молекулярного водорода, толстая многослойная оболочка, выполняющая функцию газового барьера, — все это надежно защищает азотфиксирующую систему в гетероцистах от инактивации молекулярным кислородом.
Таким образом, можно только предполагать, что механизмы нейтрализации молекулярного кислорода на различных этапах эволюции взаимодействия с ним клеток были неодинаковы. H2 каком-то этапе возникли ферментные реакции, катализирующие включение O2 в метаболизм прокариот.
МОЛЕКУЛЯРНЫЙ КИСЛОРОД В МЕТАБОЛИЗМЕ ПРОКАРИОТ
Тот факт, что все существующие на Земле прокариоты, даже строгие анаэробы, в присутствии O2 его поглощают, указывает на осуществление ими каких-то реакций взаимодействия с молекулярным кислородом. По отношению к O2 все прокариоты могут быть разделены на несколько физиологических групп (см. рис. 34). Такое подразделение говорит о необходимости или вреде молекулярного кислорода, но не раскрывает механизмов взаимодействия с ним клетки. Действительно, сейчас мы знаем, что O2 может быть необходим клетке для получения энергии или же для осуществления всего одной реакции, не имеющей энергетического значения.
| Неферментативное окисление | Ферментативное поглощение O2 (дыхание) |
| свободное окисление | окисление, сопряженное с запасанием энергии |
| нефосфорилируюшее | фосфорилирующее |
Рис. 89. Пути использования порции молекулярного кислорода, поглощенного клеткой. Объяснение см. в тексте (по Скулачеву, 1969).
H2 основании изучения энергетических процессов, происходящих в митохондриях животных клеток, В. П. Скулачев предложил следующую классификацию реакций взаимодействия клетки с молекулярным кислородом (рис. 89). Порцию поглощенного клеткой O2 можно разделить на две неравные части. Основная масса кислорода потребляется клеткой с участием клеточных ферментных систем. Поглощение клеткой какой-то части O2 не связано с ее ферментными системами. Иллюстрацией последнего служит хорошо известный факт активного поглощения кислорода суспензией убитых прогреванием клеток. В этом случае поглощение кислорода — чисто химический процесс, связанный с окислением определенных химических веществ клетки, например SH-групп клеточных белков. Нельзя исключить возможность протекания процессов аналогичной природы и в суспензии живых клеток. В свою очередь ферментативное поглощение молекулярного кислорода — дыхание64 — подразделяется на окисление, сопряженное с запасанием энергии, и свободное окисление, т. е. не связанное с запасанием энергии для клетки. Окислительные ферментативные реакции с участием O2, относимые к категории свободного окисления, — это реакции, в результате которых энергия выделяется в виде тепла65. К этой категории процессов относятся реакции, катализируемые моно- и диоксигеназами, в которых имеет место прямое включение кислорода в молекулу окисляемого вещества, а также реакции, катализируемые некоторыми оксидазами.
64 Термин "дыхание" впервые был введен для обозначения определенного процесса, связанного с жизнедеятельностью высших организмов (растений и животных). Два основных признака характеризовали этот процесс: газообмен с внешней средой с непременным участием O2; необходимость для жизнедеятельности организма. Принципиальное сходство процесса дыхания на клеточном уровне у всех высших организмов делало употребление этого термина удобным, а обозначаемое им понятие достаточно четким. Сложности возникли при применении термина "дыхание" для обозначения функционально аналогичных процессов у прокариот в силу их необычайного разнообразия. В нашем понимании термин "дыхание" распространяется на все процессы ферментативного поглощения клеткой молекулярного кислорода.
65 Реакции свободного окисления имеют важное значение в осуществлении терморегуляции у животных при охлаждении организма.
Ферментативное поглощение O2, сопряженное с запасанием энергии, подразделяется на процессы, не связанные с фосфорилированием, и процессы, сопровождающиеся фосфорилированием. В первом случае окисление, сопряженное с запасанием энергии, не связано с трансформированием свободной энергии в форму макроэргических фосфатных связей. Известно, что в клетке существуют две универсальные формы энергии: химическая и электрохимическая (H+). Один из путей получения энергии в форме трансмембранного электрохимического градиента Н+ связан с переносом электронов на O2. Энергия в этой форме может использоваться клеткой для совершения разного вида работы (см. рис. 27). Химическая энергия заключена в основном в соединениях, содержащих макроэргические фосфатные связи, и в первую, очередь в молекулах АТФ. Но на промежуточных этапах катаболических процессов, связанных в конечном итоге с поглощением O2, образуются метаболиты, содержащие богатые энергией связи, например тиоэфирные (C~S — КоА). Эти соединения могут непосредственно обеспечивать энергией некоторые биосинтетические процессы.
Наконец, при фосфорилирующем окислении энергия, высвобождаемая при электронном транспорте на молекулярный кислород и возникающая первоначально в форме H+, с помощью протонной АТФ-синтазы трансформируется в химическую форму в молекулах АТФ. В отличие от высших организмов, где достигнута высокая степень сопряжения между переносом электронов и фосфорилированием, т. е. этот путь предстает уже в сложившемся виде, у современных прокариот мы обнаруживаем различные пути переноса электронов и разные степени сопряжения электронного транспорта с фосфорилированием. Все перечисленные типы окислительных процессов с участием O2, протекающие в высокоорганизованной клетке, обнаруживаются и у прокариот.
В основу классификации, предложенной В. П. Скулачевым, положено рассмотрение всех реакций взаимодействия клетки с молекулярным кислородом под углом зрения их "энергетической значимости". По химическим механизмам, лежащим в основе этих реакций, все они могут быть разделены на 2 типа. К первому типу относятся реакции, катализируемые кислородными трансферазами, или диоксигеназами, в которых имеет место прямое присоединение молекулы кислорода к молекуле метаболита:
A + O2 AO2.
Одна молекула субстрата может акцептировать оба атома молекулы кислорода, как это имеет место в приведенной выше реакции. Акцепторами O2 могут быть молекулы двух разных субстратов:
X + Y + O2 XO + YO.
Все подобные реакции представляют собой свободное окисление и не связаны с получением клеткой энергии.
В реакциях второго типа электроны идут к кислороду, выполняющему функцию конечного акцептора. В этом случае 1, 2 или 4 электрона в зависимости от природы переносчика акцептируются молекулой кислорода, что приводит в конечном итоге к ее неполному (O2, H2O2) или полному (H2O) восстановлению. Реакции данного типа катализируются ферментами, называемыми оксидазами, и могут представлять собой свободное окисление и окисление, сопряженное с запасанием энергии. К реакциям свободного окисления относятся реакции, катализируемые растворимыми оксидазами, локализованными в цитозоле клетки. Помимо них у прокариот описан ряд связанных с мембранами оксидаз цитохромной и нецитохромной природы, перенос электронов с которых на O2 также не сопряжен с запасанием энергии.
Промежуточными по химическому механизму реакциями между приведенными выше являются реакции, в которых судьба каждого из двух атомов в молекуле кислорода различна:
A + O2 + ДH2 AO + H2O + Д.
В этом случае 1 атом поглощенной молекулы кислорода используется для окисления вещества путем прямого присоединения к нему. а другой восстанавливается до H2O в присутствии подходящего донора электронов. Обе реакции катализируются одним ферментом, принадлежащим к группе монооксигеназ, или оксигеназ (оксидаз) со смешанными функциями. Монооксигеназы в клетке многочисленны и разнообразны. Они катализируют реакции свободного окисления. Участие в процессах, сопряженных с запасанием клеткой энергии, маловероятно.
Таким образом, оксигеназы — это ферменты, катализирующие активирование O2 и последующее включение 1 или 2 его атомов в молекулы различных субстратов. Если субстратом (акцептором O2) служит водород, фермент называют оксидазой. В этом смысле оксидазы можно рассматривать как специализированный класс оксигеназ.
Оксигеназы играют важную роль в процессах биосинтеза, деградации и трансформации клеточных метаболитов: ароматических аминокислот, липидов, сахаров, порфиринов, витаминов. Субстратами, на которые воздействуют оксигеназы, часто служат сильно восстановленные не растворимые в воде соединения; их окисление приводит к тому, что продукты реакции становятся более растворимыми в воде и, следовательно, биологически активными, что важно для их последующего метаболизирования. У строго анаэробных прокариот кислород, включаемый в молекулу субстрата, происходит не из O2, а из других соединений, например воды.
Следовательно, всю совокупность взаимодействия молекулярного кислорода с клеткой, с точки зрения лежащих в основе этого химических механизмов, можно свести к участию O2 в двух типах реакций, в первом из которых он выступает в качестве конечного акцептора электронов, а во втором происходит его прямое внедрение в молекулу вещества. Только первый тип реакций с участием молекулярного кислорода может стать источником энергии для клетки. Поэтому для нас важно проанализировать эволюцию взаимодействия клетки с O2 по пути формирования ею систем, включающих использование молекулярного кислорода в качестве конечного акцептора электронов.
ФОРМИРОВАНИЕ "ОКСИДАЗНОГО МЕХАНИЗМА" ВЗАИМОДЕЙСТВИЯ С МОЛЕКУЛЯРНЫМ КИСЛОРОДОМ, СОПРЯЖЕННОГО С ЗАПАСАНИЕМ ЭНЕРГИИ
С появлением в атмосфере O2 возникла возможность переноса на него электронов. Чтобы этот перенос мог быть связан с получением энергии, необходимо было сформировать электронтранспортные цепи с определенным образом ориентированными в мембране переносчиками, обеспечивающими на отдельных этапах перемещение протонов через мембрану, а электронов — на O2, и ферментный комплекс, преобразующий возникающую при электронном транспорте электрохимическую энергию в химическую, запасаемую в молекулах АТФ.
Со сформированными электронтранспортными цепями, локализованными в мембране, содержащими все типы переносчиков и имеющими прямое отношение к получению клеткой энергии, мы уже встречаемся у рассмотренных в главах 13 и 14 анаэробных эубактерий с наиболее просто организованной энергетикой хемотрофного (брожение) и фототрофного (бескислородный фотосинтез) типа: некоторых пропионовокислых бактерий, всех фотосинтезирующих пурпурных и зеленых бактерий. В клеточных мембранах этих организмов локализованы и функционируют сопряженные с электронным транспортом АТФ-синтазы.
П. Митчелл высказал предположение, что система переноса электронов и протонов и переносящая протоны АТФаза возникли независимо друг от друга и, вероятно, неодновременно как разные способы генерации H+, необходимого для обеспечения энергией процесса избирательного транспорта питательных веществ в клетку. Последующая "встреча" обеих систем, в клетке положила начало сопряжению процессов транспорта электронов и фосфорилирования в результате обращения работы АТФазы. Это сделало возможным запасание свободной энергии окисления в молекулах АТФ. Близкий состав и аналогичная структура энергопреобразующих мембран, большое сходство механизмов сопряжения у разных групп прокариот и эукариот указывают на то, что возникшая на раннем этапе эволюции система сопряжения электронного транспорта и фосфорилирования была использована всеми организмами без принципиальных изменений.

