 2015-06-16
2015-06-16 5912
5912Способность осаждать окислы железа и марганца на поверхности клеток присуща многим эубактериям, различающимся морфологическими и физиологическими признаками и принадлежащим к разным таксономическим группам. В вопросе о том, какие организмы следует относить к железобактериям, нет единого мнения. С. Н. Виноградский впервые термин "железобактерии" применил для обозначения организмов, использующих энергию окисления Fe2+ до Fe3+ для ассимиляции CO2, т. е. способных существовать хемолитоавтотрофно. X. Молиш к железобактериям относил все организмы, откладывающие вокруг клеток окислы железа или марганца независимо от того, связан ли этот процесс с получением клеткой энергии.
Накопление окислов железа и марганца на поверхности бактериальных клеток — результат двух взаимосвязанных процессов: аккумуляции (поглощения) клетками этих металлов из раствора и окисления, сопровождающегося обильным отложением нерастворимых окислов на поверхности бактерий. Процесс аккумуляции тяжелых металлов из растворов в основе имеет физико-химическую природу и в значительной мере обусловлен химическим составом и свойствами поверхностных структур клетки. Он включает связывание металлов внеклеточными структурами (капсулы, чехлы, слизистые выделения), клеточной стенкой и ЦПМ. Сорбционные свойства поверхностных клеточных структур определяются в большой степени суммарным отрицательным зарядом молекул, входящих в их состав. Поглощение металлов приводит к значительному концентрированию их вокруг клеток по отношению к среде. Коэффициент накопления для железа и марганца может достигать величины 105–106.
Как известно, Fe2+ подвергается быстрому химическому окислению молекулярным кислородом при pH>5,5, что приводит к образованию нерастворимого Fe(OH)3. Последний вместе с Fe2+ неспецифически связывается клеточными кислыми экзополимерами. Подобный тип накопления железа не зависит от метаболической активности клеток.
Mn2+ более устойчив к окислению O2, чем Fe2+. Его химическое окисление (Mn2+ ® Mn4+) молекулярным кислородом с заметной скоростью происходит только при pH>8,5. Поэтому в нейтральной среде окисление марганца имеет только ферментативную природу. Окисление Fe2+ и Mn2+ с последующим отложением нерастворимых окислов вокруг бактериальных клеток может быть результатом взаимодействия ионов металлов с продуктами бактериального метаболизма, в частности с H2O2, образующейся в процессе окисления органических веществ при переносе электронов по дыхательной цепи. Перекись водорода, возникающая в качестве промежуточного или конечного продукта окисления, выделяется из клеток и накапливается в окружающих их структурах. В нейтральной или слабокислой среде окисление Fe2+ до Fe3+ происходит в результате непосредственного взаимодействия с H2O2:
2Fe2+ + H2O2 + 2Н+ ® 2Fe3+ + 2H2O.
Окисление марганца при взаимодействии с H2O2 осуществляется при участии каталазы, выполняющей пероксидазную функцию. Mn2+ в этом случае служит донором электронов:
Mn2+ + 2H2O2 ® MnO2 + 2H2O.
Описанные выше процессы протекают в капсулах, чехлах, слизистых выделениях, на поверхности клеточной стенки, в которых концентрируются все компоненты реакции: восстановленные формы железа и марганца, перекись водорода, каталаза. Физиологический смысл процессов окисления Fe2+ и Mn2+ с участием H2O2 — детоксикация вредного продукта метаболизма. Ни в одном случае окисление железа и марганца не приводит к получению бактериями энергии.
Наконец, среди железобактерий есть организмы, у которых окисление Fe2+ связано с получением энергии. В этом случае отложение окислов железа служит показателем активности энергетических процессов. Возможность получения энергии бактериями при окислении Mn2+ экспериментально не доказана.
В изучении железобактерий в последнее время достигнуты большие успехи, связанные с получением чистых культур ряда этих организмов. Стало понятным, что это разнообразная группа бактерий, способных окислять и откладывать окислы железа и/или марганца вне или иногда внутри клетки66. H2 основании морфологических характеристик все железобактерии могут быть разделены на две группы: нитчатые и одноклеточные.
66 К железобактериям, откладывающим железо внутри клетки, относятся магнитотактические бактерии.
К первой группе относятся грамотрицательные нитчатые бактерии, окруженные чехлом. Наиболее широко распространены представители родов Leptothrix и Sphaerotilus (см. рис. 44, 1, 2). Нити неподвижные или передвигающиеся скольжением. В чехлах, окружающих нити, накапливаются окислы железа и марганца (Leptothrix) или только железа (Sphaerotilus и др.).
Железобактерии этой группы — облигатные аэробы, но могут удовлетворительно расти при низком содержании O2 в среде. Оптимальный pH для роста — 6–8. Единственно возможный способ существования — хемоорганогетеротрофия, при этом представители рода Sphaerotilus предпочитают условия с относительно высоким содержанием органических веществ, а многие штаммы Leptothrix — среды с низким уровнем органики.
Окисление железа и марганца и отложение их окислов в чехлах этих бактерий не связано с получением ими энергии. К окислению Fe2+ при pH 6–8 могут приводить процессы как химической, так и биологической природы. Окисление марганца в этих условиях имеет биологическую природу. В обоих случаях окисление связано с действием перекиси водорода, количество которой в среде в определенных условиях может достигать 10–20 мг/л. Процесс локализован в чехлах, где концентрируются продукты метаболизма и внеклеточные ферменты. У мутантов, лишенных чехлов, накопления окислов железа и марганца не происходило. Таким образом, с помощью восстановленных форм железа и марганца обеспечивается удаление H2O2 — токсического продукта клеточного метаболизма.
Помимо бесцветных к нитчатым железобактериям относятся и некоторые фотосинтезирующие эубактерии из группы цианобактерий и скользящих зеленых бактерий.
Вторая группа железобактерий включает одноклеточные организмы из разных таксонов. Она представлена эубактериями с грамположительным и грамотрицательным строением клеточной стенки или без нее, размножающимися поперечным делением или почкованием. Клетки разной формы и размеров (форма может меняться в зависимости от стадии и условий роста), одиночные или формирующие скопления, окруженные капсулами, в которых откладываются окислы железа и марганца. Принадлежащие к этой группе железобактерии распадаются на две подгруппы, различающиеся типом метаболизма и отношением к кислотности среды.
Первая подгруппа объединяет железобактерии, растущие в. нейтральной или слабощелочной среде и характеризующиеся хемоорганогетеротрофным типом метаболизма. Представители подгруппы — свободноживущие микоплазмы, объединенные в роды Metallogenium, Gallionella, Siderococcus. Им свойствен типичный для микоплазм полиморфизм: кокковидные клетки, от которых могут отходить тонкие нити, пучки переплетенных тонких нитей и т. д. H2 поверхности нитей откладываются окислы железа (Gallionella, Siderococcus) или железа и марганца (Metallogenium). Растут в нейтральной или кислой среде. Некоторые из них олиготрофы. Все — аэробы или микроаэрофилы. Отложение окислов железа и марганца — результат химических реакций или функционирования перекисного пути и не имеет отношения к получению клетками энергии.
Вторую подгруппу составляют в большинстве аэробные ацидофильные формы. Оптимальный pH их роста лежит ниже 4,5 (2–3). В этих условиях Fe2+ в присутствии O2 устойчиво к химическому окислению. Для ацидофильных железобактерий установлена способность получать энергию в результате окисления двухвалентного железа.
Основным представителем железобактерий с энергетическим метаболизмом хемолитотрофного типа является Thiobacillus ferrooxidans, относящийся к группе тионовых бактерий и обладающий способностью получать энергию также в результате окисления различных восстановленных соединений серы. Окислять закисное железо с получением клеткой энергии способна и выделенная недавно облигатно ацидофильная бактерия Leptospirillum ferrooxidans, близкая по ряду свойств к Т. ferrooxidans, но в отличие от последнего не окисляющая соединения серы.
Leptospirillum ferrooxidans и большинство изученных штаммов Т. ferrooxidans принадлежат к облигатным хемолитоавтотрофам, использующим энергию окисления железа для ассимиляции CO2, служащей основным или единственным источником углерода. Некоторые штаммы Т. ferrooxidans оказались способными расти на средах с органическими соединениями, являясь, таким образом, факультативными хемолитоавтотрофами. Наконец, описаны термофильные бактерии, получающие энергию в результате окисления Fe2+ и нуждающиеся для роста в органических соединениях, т. е. осуществляющие метаболизм хемолитогетеротрофного типа.
Окисление железа, приводящее к получению энергии, происходит в соответствии с уравнением
2Fe2+ + 1/2O2 + 2Н+ ® 2Fe3+ + H2O,
что сопровождается незначительным изменением уровня свободной энергии (D G 0' при pH 2 равно — 33 кДж/моль). Поэтому для обеспечения энергией клетке необходимо "переработать" большие количества железа.
Механизм окисления Fe2+ в дыхательной цепи изучен у Т. ferrooxidans. Дыхательная цепь этой бактерии содержит все типы переносчиков, характерные для дыхательной системы аэробных хемоорганотрофных эубактерий, но участок цепи, связанный с получением энергии, очень короток (рис. 98, А).
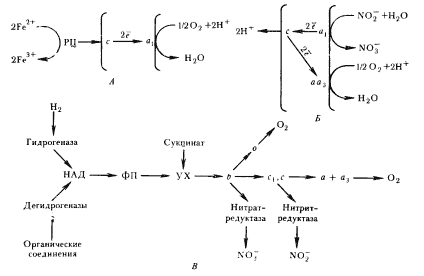 |
| Рис. 98. Схема энергетических процессов у представителей разных групп хемолитотрофных эубактерий: А — Thiobacillus ferrooxidans; Б — Nitrobacter; В — Paracoccus denitrificans: РЦ — рустицианин; ФП — флавопротеин; УХ — убихинон; b, c, o, a1, aa3 — цитохромы (по Haddock, Jones, 1977; Jones, 1980) |
Окисление Fe2+ происходит на внешней стороне ЦПМ; в цитозоль через мембрану железо не проникает. Электроны с Fe2+ акцептируются особым медьсодержащим белком — рустицианином, находящимся в периплазматическом пространстве.
Затем с рустицианина они передаются на цитохром c, локализованный на внешней стороне ЦПМ, а с него на цитохром a1, расположенный на внутренней стороне мембраны. Перенос электронов с цитохрома a1 на 1/2O2, сопровождающийся поглощением из цитоплазмы 2H+, приводит к восстановлению молекулярного кислорода до H2O. Особенность дыхательной цепи Т. ferrooxidans — отсутствие переноса через мембрану протонов, а перенос только электронов. Градиент H+ по обе стороны ЦПМ поддерживается как за счет поглощения протонов из цитоплазмы, так и в результате низкого pH внешней среды, в которой обитают эти бактерии. Синтез АТФ происходит за счет движения H+ из внешней среды в цитоплазму через АТФ-синтазный комплекс. Движущей силой служит в основном DpH. Для синтеза 1 молекулы АТФ необходимо окислить как минимум 2 молекулы Fe2+.
Образование восстановителя происходит в результате энергозависимого обратного переноса электронов. Активность участка дыхательной цепи, обеспечивающей обратный электронный транспорт, на порядок ниже активности короткого участка, функционирование которого приводит к получению энергии. В целом для фиксации 1 молекулы CO2 в восстановительном пентозофосфатном цикле необходимо окислить больше 22 молекул Fe2+. Таким образом, из всех представителей эубактерий, у которых обнаружена способность к окислению железа и/или марганца, только облигатно ацидофильные формы могут использовать энергию окисления Fe2+ для ассимиляции CO2, т. е. существовать хемолитоавтотрофно. Именно они являются истинными железобактериями, соответствуя тому названию, которое было введено С. Н. Виноградским.
Для остальных организмов образование окислов железа и марганца не связано с получением энергии и происходит в результате неспецифических реакций ионов металлов с продуктами метаболизма, прежде всего продуктами неполного восстановления O2. Неспецифичность функции перекисного окисления железа и марганца, проявляющейся у широкого круга эубактерий, ставит вопрос о правомерности использования термина "железобактерии" в значении, предложенном X. Молишем. Некоторые авторы в связи с этим считают целесообразным для обозначения остальных организмов использовать названия "железоокисляющие" и "марганецокисляющие" бактерии.
Эубактерии, описанные в этом разделе, широко распространены в природе и могут существовать в большом диапазоне условий. Облигатные ацидофилы обнаружены в подземных водах сульфидных месторождений, кислых водах железистых источников и кислых озерах с высоким содержанием закисного железа. Нитчатые формы также занимают вполне определенные экологические ниши. Представители рода Leptothrix — обитатели олиготрофных железистых поверхностных вод, Sphaerotilus предпочитают среды с высоким содержанием органических веществ.








