 2015-06-16
2015-06-16 2444
2444Уксуснокислые бактерии, выделенные в роды Gluconobacter и Acetobacter, могут получать энергию, осуществляя неполное окисление ряда органических соединений. Это грамотрицательные бесспоровые палочки, слабоподвижные за счет перитрихиально или полярно расположенных жгутиков, или неподвижные. Облигатные аэробы. Довольно требовательны к субстратам для роста. Почти все виды нуждаются в отдельных витаминах, в первую очередь в пантотеновой кислоте, однако есть формы, способные к синтезу всех факторов роста.
К числу окисляемых соединений относятся одноатомные спирты, содержащие от 2 до 5 углеродных атомов, а также многоатомные спирты — производные сахаров. Окисление первичных спиртов приводит к образованию кислот. Например, этанол с помощью соответствующих дегидрогеназ окисляется до ацетата:
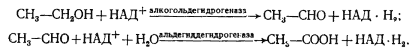
Вторичные спирты окисляются до кетонов:
CH3-CHOH-CH3 ® CH3-CO-CH3.
Многоатомные спирты окисляются этими бактериями в альдозы и кетозы, например: сорбит ® сорбоза; глицерин ® диоксиацетон. Альдозы и кетозы могут далее окисляться в соответствующие кислоты. Метаболизирование cахаров осуществляется по окислительному пентозофосфатному пути.
Круг окисляемых соединений различен для разных представителей, входящих в эту группу. С точки зрения характеристики энергетических возможностей уксуснокислых бактерий важно подчеркнуть, что у них развилась удивительная способность воздействовать на определенные химические группировки" осуществляя их одно- или двухступенчатое окисление. Наиболее характерна способность этих бактерий окислять этиловый спирт в уксусную кислоту, давшая название всей группе в целом.
Дальнейшая судьба полученных в результате неполного окисления продуктов различна. Некоторые уксуснокислые бактерии не способны к последующим превращениям образовавшихся соединений, и их окислительные способности, следовательно, весьма ограничены. Эти бактерии, объединенные в род Gluconobacter (единственный вид G. oxydans), глюкозу окисляют до глюконовой кислоты, этанол — только до ацетата, который дальше не может ими окисляться из-за отсутствия "замкнутого" ЦТК.
Вторую группу составляют бактерии, способные к полному окислению органических субстратов до CO2 и H2O. В этом случае образовавшаяся уксусная кислота представляет собой лишь промежуточный этап, и после исчерпания из среды исходного субстрата бактерии начинают медленно окислять уксусную кислоту, включая ее в механизм конечного окисления — ЦТК. Бактерии этой группы объединены в род Acetobacter, типичным представителем которого является A. peroxydans.
Электроны от окисляемых субстратов поступают в дыхательную цепь и далее через систему переносчиков передаются на O2, служащий обязательным конечным акцептором электронов. Электронный транспорт приводит к генерированию DmH+.
Место включения электронов в дыхательную цепь определяется ферментом, катализирующим соответствующую окислительную реакцию. Если окисление катализируется НАД-зависимой дегидрогеназой, электроны (водород) передаются на НАД+ и с него на переносчики, локализованные на мембране, что открывает возможность для сопряжения электронного транспорта с тремя трансмембранными перемещениями протонов и, соответственно, синтезом 3 молекул АТФ. Недавно у представителей родов Acetobacter и Gluconobacter были обнаружены дегидрогеназы, содержащие в качестве простетической группы соединение из группы хинонов, способные принимать и отдавать 2 атома водорода. Хинонсодержащие дегидрогеназы локализованы на внешней стороне ЦПМ, где и происходит окисление этанола и других соединений. Электроны поступают в дыхательную цепь на уровне цитохромов, а протоны выделяются в периплазматическое пространство.
Уксуснокислые бактерии часто развиваются вслед за дрожжами, используя продукт спиртового брожения как субстрат для роста. Применяются в микробиологической промышленности для получения столового уксуса и в производстве аскорбиновой кислоты (на этапе окисления сорбита в сорбозу).

