 2013-12-31
2013-12-31 912
912ГЕНЕТИЧЕСКАЯ ИЗМЕНЧИВОСТЬ ФИТОПАТОГЕННЫХ
Она происходит путем:
• наследуемых изменений, приводящих к появлению новых признаков за счет изменения последовательности ДНК — мутации;
• обмена участками ДНК — рекомбинации внутри генома;
• обмена генетической информацией между бактериями — трансдукции, конъюгации и трансформации (рис. 38).
Мутации. Скачкообразные наследуемые изменения, приводящие к появлению новых признаков, называются мутациями. В большинстве своем они приводят к исчезновению признака и снижению жизнеспособности, реже — к появлению нового признака, придающего организму новые свойства. Наиболее важные мутации у фитопатогенных бактерий происходят в генах авирулентности (avr) и генах группы сверхчувствительности (hrp), приводя к изменению расовой принадлежности и снижению или усилению агрессивности патогена. Например, у возбудителя черной бактериальной пятнистости томата и перца (Xanthomonas campestris pv. vesicatoria) и возбудителя сосудистого бактериоза капусты (X. campestris pv. campestris) замена нуклеотидов или удлинение пятинуклеотидного повтора в гене avrBs2 приводит к появлению но-
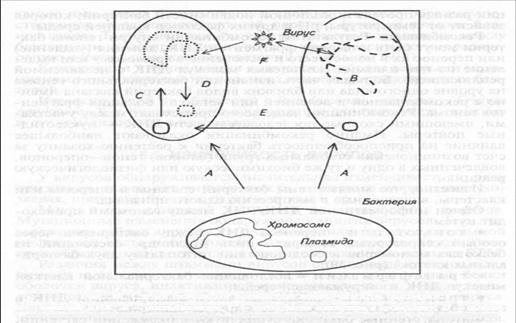
Рис. 38. Схема основных процессов изменчивости бактерий:
А — мутация ДНК; В — рекомбинация; С — интеграция плазмиды в хромосому; D — выделение части хромосомы в плазмиду; Е — конъюгация или трансформация; / — трансдукция
вых рас, поражающих растения с генами устойчивости Bs2 (перец) или Rxcl (абиссинская капуста). Изменение числа повторов нуклеотидов в середине гена avrBs3/Pth у Х.с. pv. vesicatoria и возбудителя бактериального рака цитрусовых X. citri также приводит к смене расы патогена.
Выявлено, что высокая нестабильность геномов фитопатогенных бактерий зачастую связана с активностью мобильных генетических элементов (транспозонов), фланкирующих гены, которые участвуют в патогенезе. Так, у Pseudomonas syringae гены авирулен-тности инактивируются вставкой мобильного элемента при заражении устойчивого растения, что также приводит к восстановлению вирулентности бактерии.
Различия в агрессивности бактерий и симптомах вызываемых ими болезней иногда определяются мутациями отдельных генов или изменением их регуляции. Например, генетические различия между ксантомонадами, вызывающими листовую пятнистость и сосудистый бактериоз у крестоцветных, заключаются в активизации разных протеаз и различной подвижности бактерий, которая зависит от температуры, рН и других факторов внешней среды.
Рекомбинации. Результатом рекомбинации внутри генома бактерии могут быть инверсия фрагментов ДНК, делеция (удаление) или перенос их в новое место и включение в хромосому или выделение из нее плазмид (кольцевых молекул ДНК с независимой репликацией). Большая часть изменений бактериального генома на уровне одного вида или близких видов бактерий связана именно с рекомбинацией и делецией или вставкой больших фрагментов генома. Рекомбинации чаще всего происходят между участками, имеющими сходные последовательности ДНК — нуклеотидные повторы. Именно рекомбинации оказывают наибольшее влияние на приспособленность бактерии к растению-хозяину за счет возникновения стабильных групп (блоков) генов—оперонов, вовлеченных в одну и ту же биохимическую или физиологическую реакцию.
Известно, что многие гены бактерий связаны в опероны или кластеры, вовлеченные в экспрессию одного признака.
Обмен информативной ДНК/РНК между геномами происходит путем:
• конъюгации —переноса ДНК между бактериями через особый секреторный аппарат — пиль (цилиндр, состоящий из белковых мономеров и соединяющий цитоплазму двух бактериальных клеток) (рис. 39);
• трансформации — поглощения бактериальной клеткой молекул ДНК из окружающей среды;
• трансдукции — внедрения части бактериальной ДНК в геном бактериофага и перенос ее в другую бактерию.
Фитопатогенные бактерии существуют в окружении растений, заселенных другими бактериями, что может привести к приобретению ими новых биохимических свойств и нового образа существования через горизонтальный перенос функциональных блоков генов. Такой перенос генов при обмене плазмидами между бактериями часто приводит к увеличению агрессивности и расширению спектра поражаемых растений (Gunsalus, 1978).
Конъюгация и трансдукция являются наиболее важными механизмами горизонтального переноса генов у фитопатогенных бактерий.
Известно, что у бактерий Pseudomonas syringae, Erwinia amylovora, X. с. pv. vesicatoria и др. важные для патогенеза гены hrp и avr локализованы в плазмидах.









