 2015-06-16
2015-06-16 3704
3704Если мы сравним, например, значение орнитологии и множества работ учёных, изучающих около одного килограмма птиц на гектаре, с чрезвычайно ограниченным числом работ исследователей, интересующихся сотнями килограммов или тоннами дождевых червей на гектаре, мы должны заключить, что наши знания об экосистемах сильно искажены знанием природы, визуально воспринимаемой выше уровня земли, и игнорированием жизни под землёй.
Буше, М.Б., (1985)
Большинство дождевых червей - почвенные организмы, однако некоторые виды, например Pontodrilus bemiudensis, отлично чувствуют себя в эстуариальных водах. Несмотря на то, что для таксономической классификации желательно идентифицировать каждый вид дождевых червей, классификация, основанная на экологической стратегии, более подходит при использовании дождевых червей в практической деятельности человека. Даже такая классификация может быть несостоятельной, как, например, в условиях тропического климата, где эти черви могут быть найдены под влажными, гниющими брёвнами, в рисовых чеках, в дамбах, на террасах, и даже под обычными домашними жерновами там, где в результате постоянного использования скапливаются значительные количества органического вещества.
В мире имеется пять обширных таксономических семейств дождевых червей: Moniligastridae, Megascolecidae, Eudrilidae, Giossoscolecidae и Lumbricidae (Jamieson, 1971). Julka (1988) описал индийские виды олигохет или малощетинковых в монографии «Фауна Индии». Таксономические исследования дождевых червей Индии проводились в основном доктором G. E. Gates, который является основателем современной таксономии дождевых червей. Однако, много таксономических работ по индийским видам было выполнено Julka (1975, 1976, 1977, 1978, 1979,1981 и 1983); Jamieson (1977) и Easton (1982).
КЛАССИФИКАЦИЯ ДОЖДЕВЫХ ЧЕРВЕЙ ПО ТИПУ ПИТАНИЯ
Дождевые черви являются сапрофагами и на основании особенностей их способов питания они могут быть подразделены на две группы: детритофаги и геофаги (Lee, 1985).
Детритофаги кормятся на поверхности или близ поверхности почвы. Они питаются в обогащенном органикой почвенном горизонте, поедая в основном остатки растений и отмершие корни или экскременты млекопитающих. Таких червей называют создателями гумуса, они включают в себя две формы: эпигеики и анецики(см. ниже). Такие виды, как Perionyx excavatus (фото 1), Eisenia foetida, Eudrilus euginiae, Lampito mauritii (фото 2), Polypheretima elongata, Octochaetona serrata (фото 4) и Octochaetona surensis являются только некоторыми представителями питающихся детритом дождевых червей.
Черви-геофаги, обитают и питаются глубоко под поверхностью земли, поглощая большие количества почвы, богатой органикой. Их обычно называют потребителями гумуса, и они представляют собой дождевых червей эндогеиков. Два типичных представителя таких червей - Octochaetona thurstoni (фото 3) и Metaphire posthuma.
Буше (1977) разделил дождевых червей на три типа: epigeics, anecics и endogeics или эпигеики, анецики, и эндогеики (рис. 1).
Эпигеики – поверхностнороющие виды дождевых червей, эффективно размельчающие и фрагментирующие лиственный опад. Они являются фитофагами. Обычно они не влияют на почвенную структуру, поскольку не могут жить в почве.
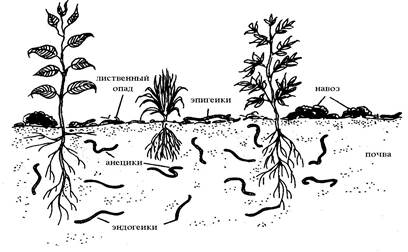
Рис. 1. Классификация дождевых червей, основанная на экологической стратегии.
Анецики поглощают лиственный опад, перемешанный с почвой верхних слоёв, и поэтому должны называться геофитофагами. Они так же могут производить на поверхности почвы копролиты, обычно в зависимости от ее плотности.
Эндогеики являются геофагами. Они живут глубоко в почве, получая питательные вещества из поглощаемой ими почвы, богатой органикой.
Различные по поведению, морфологии и физиологии обитающие в почве непигментированные формы и обитающие на поверхности почвы сильнопигментированные формы дождевых червей представляют собой эволюционные полюса, возникшие в результате r- и K- отбора при содействии адаптивной радиации (Satchell, 1980).
ДРИЛОСФЕРА
Дрилосфера включает в себя собственно самих дождевых червей и общий объём почвы и опада, на который они влияют через свою активность, в мутуалистической связи со свободными почвенными микроорганизмами, и, возможно, некоторыми облигатными симбиотическими простейшими или бактериями (Rouelle с сотр., 1985; Lavelle и Martin 1992). Дождевые черви - анецики и эндогеики, связанные со свободноживущими почвенными бактериями, образуют дрилосферу (рис. 2).
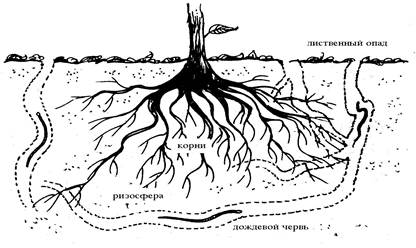
Рис. 2. Дрилосфера.
Было установлено, что почва, поглощенная и переваренная дождевыми червями, вместе с выделяемой из кишечника слизью, стимулирует микробиологическую активность. Созданные структуры и ходы, особенно вертикальные ходы червей-анециков и горизонтальные ходы червей-эндогеиков, усиливают проницаемость и циркулирование воды и газов в почве, что дополнительно ускоряет разложение органических веществ (Tisdal и Oades, 1982).
ФИЗИЧЕСКИЕ ВОЗДЕЙСТВИЯ ДОЖДЕВЫХ ЧЕРВЕЙ
НА ПОЧВУ
Дождевые черви-анецики, которые преимущественно ответственны за создание вертикальных ходов, оставляют свои копролиты, в основном, на поверхности почвы, особенно на твёрдой (Habibullah и Ismail, 1985; Krishnamoorthy, 1989), в то время как дождевые черви-эндогеики, обычно делающие горизонтальные ходы, оставляют свои копролиты внутри ходов. Интересно заметить, что поверхностные копролиты играют значительную роль в развитии почвенного профиля, тогда как копролиты, находящиеся под поверхностью, участвуют в педогенезе. Известно, что дождевые черви измельчают почву (Evans, 1948; Teotia с сотр., 1950; Shrikande и Pathak, 1951; Joshi и Kelkar, 1952). Однако, принимая во внимание то, что почва в кишечнике самого дождевого червя находится относительно непродолжительное время, не похоже, что подобное измельчение играет значительную роль (Lee, 1985). Копролиты образуют почвенные агрегаты (они стабилизируются после их выделения), которые склеиваются в результате микробиологического (ферментатитвного) расщепления входящих в их состав органических компонентов (Waksman и Martin, 1939) или связывания гифами грибов (Parle, 1963b). Количество поверхностных копролитов, выделяемых дождевыми червями, варьирует в зависимости от внешних условий и составляет от 0,25 кг/м2 на австралийских пастбищах (Barley, 1959) до 25,75 кг/м2 на немецких пастбищах (Graft, 1971).
Дождевые черви сильно изменяют пористость почвы. Еще Wollny в1890 году сообщал, что в почве, заселённой дождевыми червями, возрастает величина соотношения объемов почва/воздух от 8 до 30 %. В связи с этим дождевые черви так же влияют на проникновение воды и ее водоудерживающую способность почвы. Данные Stockdill (1966)) относятся к доступной влаге в почве (0-30 см) под участками пастбищ в присутствии или отсутствии люмбрицид в Новой Зеландии. Было поставлено несколько экспериментов для изучения влияния червей на физические свойства почв, особенно относительно подвижности воды. Несмотря на множество трудностей в осуществлении этих опытов, они были необходимы и дали полезную информацию для определения функционально важных морфологических изменений (Diestel, 1992). Кроме дождевых червей-анециков Lumbricus terrestris (Joschko с сотр., 1989), виды-эндогеики Apporrectoidea caliginosa и Apporrectoidea rosea (Joschko с сотр., 1992) так же создают водопроводящие каналы, которые эффективны в насыщенных или затопленных водой почвах. Физические свойства почв так же влияют на биологическую активность. Такие факторы как перенос воздуха, воды и тепла определяются топологией и пористостью системы (Kretzschmar и Monesriez, 1992).
Известно, что вытаптывание влияет на физические и химические свойства почв (Briggs, 1978), природу и биоразнообразие флоры и фауны, что в конечном счёте приводит к снижению биомассы дождевых червей (Chappell с сотр., 1971; Aritajat с сотр., 1977; Briggs, 1978; Piearce, 1984; Piearce с сотр., 1995). Хотя вытаптывание сильно влияет на численность поверхностно обитающих видов дождевых червей, глубоко обитающие виды, например Apporrectoidea longa, являются частично устойчивыми.
Надежда на поддержание местного видового разнообразия дождевых червей-анециков в сложных и трудных природных условиях реализуется на примере дождевого червя вида L. mauritii. Этот вид не только легко поддерживать, но так же его можно разводить на песках и песчаных почвах, переселяя этих дождевых червей из легко доступных местных источников. Такие вермикультуры не только могут служить in situ, то естьна месте в качестве почвоулучшителей, но и поддерживать рост самих растений (Anne Grace и Ismail, 1994).
ХИМИЧЕСКИЕ ВОЗДЕЙСТВИЯ ДОЖДЕВЫХ ЧЕРВЕЙ НА ПОЧВУ
Поскольку дождевые черви перерабатывают большие количества поверхностного опада и других органических материалов, они влияют на химический состав почв, а так же на транспорт питательных веществ и, таким образом, на доставку их растениям. Дождевые черви осуществляют транспорт органического материала в низлежащие почвенные слои, при этом большая часть материала переваривается, мацерируется и снова выделяется. Сами дождевые черви так же вносят в почву питательные вещества в виде азотсодержащих отходов их жизнедеятельности. Аммиак, который составляет значительную часть азотсодержащих веществ, выделяемых дождевыми червями (Cohen and Lewis, 1949), может вызвать временное увеличение значения рН почвы. Однако, дождевые черви более чувствительны к компонентам, ответственных за создаваемое конкретное значение рН окружающей среды, чем к значению рН, как таковому (Magdoom и Ismail, 1986).
Некоторые исследователи сравнивали химический состав копролитов дождевых червей с таковым окружающей их почвой. Их результаты были сравнимы с данными, полученными ранее Dusserre (1902), и однозначно показали, что копролиты дождевых червей имеют более высокую общую обменную ёмкость, и, в основном, они богаче по содержанию общего органического вещества, фосфора, обменного калия и магния, а также общего обменного кальция. Дождевые черви, однако, похоже способствуют нитрификации (Keller, 1893; Puh, 1941), поскольку они способствуют росту бактериальных популяций (Parle, 1963 a,b) и аэрации почвы (Heymons, 1923; Ansett, 1951). Несмотря на то, что при прохождении через кишечник дождевого червя популяция азотобактера может уменьшаться (Ruschmann, 1953, 1954), она может значительно увеличиваться затем в самих выделенных копролитах (Stockli, 1928). Поскольку в тканях дождевых червей содержится много белка (Alawdeen и Ismail, 1986), при разложении его так же образуется нитратный азот (Russell, 1910).
Дождевые черви поглощают не все растительные остатки, экскременты и другие органические вещества, которые они перемещают с поверхности почвы. Фактически из заглатываемого количества корма переваривается лишь небольшая ее часть (Lee, 1985). Заглатываемый органический материал мацерируется, перемешивается с поглощённым неорганическим материалом, проходит через кишечник и выделяется в виде копролитов, которые имеют небольшие химические изменения, но у них существенно изменяется физическая структура. Прежде всего, увеличивается общая поверхностная площадь, что также повышает микробиологическую активность. Таким образом, наиболее важная функция дождевых червей – стимуляция активности микроорганизмов, которая происходит в копролитах (Nowak, 1975); при этом усиливается трансформация растворимого азота в микробный белок, что предотвращает его потери от вымывания в нижние горизонты почвы. В дрилосфере присутствуют 40% всех аэробных фиксаторов азота, 13% анаэробных фиксаторов и 16% денитрификаторов; максимальная плотность популяций наблюдается в слое почвы от 20 до 40 см (Bhatnagar, 1975). Величина отношения C:N в копролитах дождевых червей однако несколько выше, чем таковая в окружающей почве (Graff, 1971; Lavelle, 1978), при этом количества углерода и азота, выделяемые в копролиты, иногда составляют довольно большую величину (Syers с сотр., 1979).
Lee (1983) суммировал данные о влиянии дождевых червей на почвенный азот и его круговорот. В соответствии с его выводами, азотсодержащие продукты метаболизма дождевых червей возвращаются в почву через копролиты, мочу, мукопротеины и отмершие ткани червей. Только поступление азота в почву в виде тканей умерших червей может составлять около 6-7 г/ярд2 (Satchell, 1963); а при выделении мочи и слизи – до 10 г/ярд2 (Needham, 1957). Другими источниками поступления азота в почву являются разлагающиеся корни, выделения беспозвоночной и позвоночной фауны, отмершие растительные и животные материалы и микробиологическая азотфиксация. По данным Scheu (1994), в почве существует мобилизуемый дождевыми червями азотный пул, связанный с мобилизуемыми дождевыми червями источниками углерода, и этот физически защищенный азот и углерод в значительной степени пополняет мобилизуемые дождевыми червями азотный и углеродный пулы.
ВЛИЯНИЕ ОТСУТСТВИЯ ДОЖДЕВЫХ ЧЕРВЕЙ НА ПОЧВУ
Очень большие количества невозобновляемого сырья расходуется на производство химических удобрений и биоцидов, которые поступают в почву, через почву затем в растения и далее по всей пищевой цепи. Первоначально это положительно влияло на уровень сельскохозяйственного производства, но эта длительная практика привела к деградации почв и исчезновению из неё жизни. Фраза: «Мы не унаследовали землю от наших предков, а заняли её у наших детей» останется горьким упреком всем нам, если мы не сделаем что-нибудь для спасения умирающей почвы. Почва является «живой» только тогда, когда в ней есть обитатели в виде флоры и фауны.
Инсектициды вызывают устойчивость. Для того, что бы выиграть войну с насекомыми люди должны научиться не распылять их (Ffrench-Constant, 1995). Однако вопрос состоит в том, кто будет учить тому, чтобы не распылять их? Средний фермер в Индии (маргиналы или чернорабочие на больших фермах) распыляет инсектициды, даже не используя защитные средства. Такое распыление в различных формах пагубно влияет на все организмы – от микробов до человека. Они так же вызывают гибель и хищников, и полезных организмов, например пауков, ос и других животных, питающихся вредными насекомыми.
Глава 3.
Дождевые черви: структура.
В студенческой аудитории дождевой червь типа Аннелида знаком каждому студенту-биологу. В зоологических руководствах описаны его анатомические особенности, а множество материалов отмечают поразительные формы и функцию дождевых червей. Но наряду с этим студенты и преподаватели сталкиваются с массой публикаций, спектр которых широк по содержанию: от военной ценности дождевых червей как пищи для выживания и до того, что их впрок заготавливают кроты. Такое количество литературы довольно необычно о группе беспозвоночных животных, которые не имеют важного значения ни как паразиты, ни как источник пищи. Это связано с зоологическим интересом к дождевым червям, как наземным организмам, которые унаследовали многие признаки от своих водных предков; от того, что дождевые черви являются удобной экспериментальной моделью для физиологов и этологов, до того, что они влияют на почвенное плодородие.
Satchell, J.E., (1972)

