 2015-06-26
2015-06-26 1270
1270Первичные повреждения,
вызываемые в ДНК ионизирующей радиацией
Большинство механизмов повреждающего действия облучения на биомолекулы ДНК изучены в модельных экспериментах. Однако существующие литературные данные позволяют представить некоторую схему событий, происходящих в облученной ДНК.
На первом этапе облучения в хромосомной ДНК образуется сложный макрорадикал с двумя типами локализации неспаренных электронов, которым соответствуют повреждения оснований (потеря и химические модификации) и повреждения сахарофосфатных цепей (одиночные и двойные разрывы фосфодиэфирных и межуглеродных связей). Причем нуклеотидные основания повреждаются чаще, чем сахарофосфатная часть ДНК, примерно в 3 раза [Шальнов М.И.,1972г.,1973г.]. По данным работы [Шариатый В.А., 1972г.], 80-90% электронов и радикалов, образующихся при облучении, реагируют с азотистыми основаниями – самыми лабильными частями молекулы – и только 10-20% энергии расходуется на разложение сахарофосфатного фрагмента ДНК. Наиболее радиоустойчивыми являются углеродные связи, которые в 7,5 раза стабильнее, чем фосфодиэфирные. Из азотистых оснований большей радиочувствительностью обладают пиримидиновые, из которых самый лабильный тимин. Пуриновые основания в 2 раза устойчивее пиримидиновых.
Под действием ионизирующей радиации чаще повреждается одна нить ДНК. При этом образуются такие первичные повреждения, как дезаминирование оснований, алкилирование, возникновение оксимов цитозина, димеризация или гидратация пиримидиновых оснований и др. Вторичные реакции – это разрывы водородных связей, конфигурационные изменения надмолекулярных структур ДНК, а также внутри- и межмолекулярные сшивки полимерных цепей. Разрушение надмолекулярной структуры (деспирализация) обусловлено двойными разрывами ДНК, которые возникают, если одиночные разрывы нитей ДНК расположены на расстоянии не более 10 пар нуклеотидов друг от друга [Шальнов М.И., 1973г.].
Попадание кванта энергии при облучении клетки – процесс случайный, однако конечное повреждение образуется преимущественно в определенных участках хромосом, в так называемых „горячих точках”. Это явление можно объяснить миграцией энергии или заряда вдоль хромосомы к наиболее „ранимым” местам. Существует миграция электронного возбуждения по молекуле ДНК на большие расстояния – 103-104 пар оснований [Шафрановская Н.И. и др., 1972г.]. Установлено, что если первоначально поражается наиболее радиоустойчивая компонента в смеси „фосфат-сахар” или в сахарофосфате, то, в конечном счете, свободная валентность локализуется на сахарной компоненте этой смеси [Шариатый В.А., 1972г.]. Далее миграция энергии или заряда идет в направлении наиболее легко возбуждаемых азотистых оснований, которые и являются преимущественными местами атаки электронов. С сахарным фрагментом ДНК электрон практически не реагирует, а лишь стабилизируется и за счет этого принимает участие в реакциях с азотистыми основаниями ДНК [Шарпатый В.А., 1972г.]. По данным работы [Gregoli S и др., 1972г.], образующийся под действием γ-облучения анион-радикал может возникать случайно на любом из четырех нуклеотидов, а затем неспаренный электрон передается на тот нуклеотид, который обладает наибольшим электронным сродством. После этого анион-радикал превращается в стабильный. Пиримидиновые основания как наиболее легко возбуждаемые и являются, по мнению авторов, точками приложения повреждающего действия радиации. С пуриновых же оснований сигнал ЭПР полностью снимается. Таким образом, независимо от места первоначального попадания кванта энергии первичные повреждения начинают формироваться на пиримидиновых нуклеотидах. Более того, миграция энергии и заряда возможна не только внутри молекулы ДНК, но и с ДНК на протектор. При введении, например, профлавина – лучшего, чем любой нуклеотид, акцептора электронов – конечное радиационное повреждение локализуется на профлавине. Если же ввести гистамин, который является худшим акцептором электронов, чем азотистые основания, то после γ-облучения сигнал передается, наоборот, от гистамина к нуклеотидам [Gregoli S и др., 1972г.].
Вещества-радиопротекторы могут захватывать на себя и электроны и возбуждение, чем вызывают снижение выхода радикалов ДНК [Шарпатый В.А., 1972г.]. Предполагается, что такой радиопротектор образует с ДНК комплекс, в котором возможна миграция заряда или возбуждения с ДНК на защитное вещество, а рекомбинация этого заряда с зарядом противоположного знака происходит на фрагменте протектора.
Таким образом, если квант энергии попадает на ДНК, необязательно последует ее повреждение: при наличии в клетке веществ-радиопротекторов возможна миграция энергии или заряда полностью или частично на протектор (рис. 1.1). Если такой миграции нет, то первичное радиационное повреждение локализуется на азотистых основаниях ДНК.
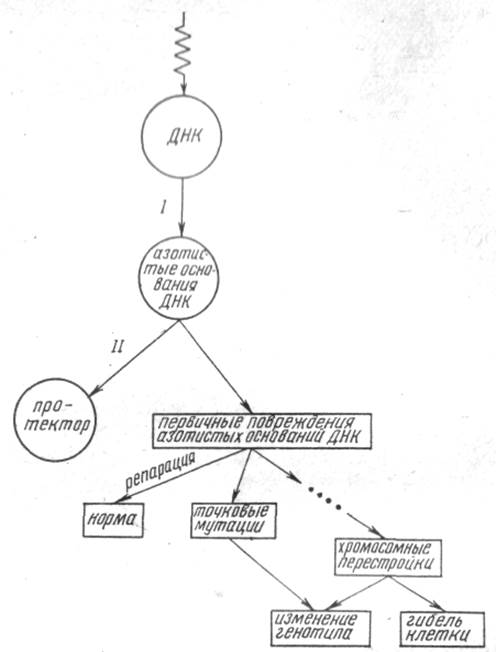
Рис. 1.1. Схема радиогенетических эффектов в клетке эукариот
Какова дальнейшая судьба этих повреждений? Во-первых, первичные повреждения могут быть репарированы; во-вторых, возможно образование точковых мутаций и, в-третьих, первичные радиационные повреждения нуклеотидов могут дать начало цепи событий, приводящих к формированию хромосомных аберраций (см. рис. 1.1).
Поскольку репарация радиационных повреждений ДНК описана во многих работах, остановимся на вопросах образования точковых и хромосомных мутаций.








