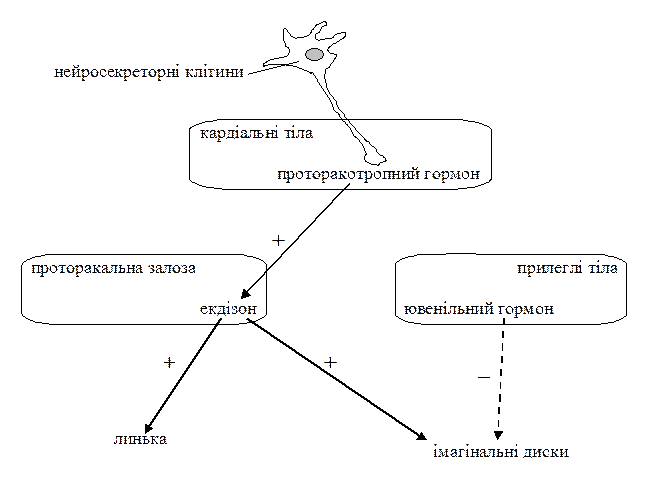2015-06-28
2015-06-28 62457
62457Автор: преподаватель, Варенюк И. М., кандидат биологических наук, доцент. Тип материала: Учебное пособие
Прямий розвиток зустрічається у безхребетних (вільноживучі плоскі черви, коловертки, малощетинкові черви (oligocheta), п’явки, павукоподібні) та хордових (круглороті (міксини), частина риб, плазуни, птахи, ссавці). При цьому з яйцевих оболонок або організму матері виходить (народжується, вилупляється) особина, зовні схожа на дорослий організм. Відмінності стосуються, головним чином, розмірів тіла, деяких пропорцій, недорозвитку деяких органів та систем органів, нездатністю до розмноження (недорозвинута статева система).
Окрім прямого розвитку, існує ще й непрямий розвиток, або розвиток з метаморфозом. Цей тип розвитку широко поширений серед безхребетних, є він і у нижчих хребетних (асцидій, частини риб, амфібій). Серед безхребетних розвиток з метаморфозом зустрічається у губок, кишковопорожнинних, паразитичних плоских червів, немертин, нематод, багатощетинкових червів (polychaeta), більшості ракоподібних, комах, молюсків, напівхордових, голкошкірих.
При такому типі розвитку з яйця виходить личинка, зовні не схожа на дорослу особину. Після певного періоду личинка починає перетворюватися на дорослу особину, цей процес називається метаморфозом. Є декілька типів метаморфозу: еволютивний (перетворення личинки на дорослу особину відбувається поступово) (наприклад, кільчасті черви, ракоподібні), революційний (катастрофічний) (відбувається швидке перетворення личинки на дорослу особину) (наприклад, комахи з повним перетворенням), некробіотичний (під час метаморфозу дегенеративні зміни переважають над прогресивними) (наприклад, у асцидій). Крім того, метаморфоз поділяють на первинний і вторинний (наприклад, у комах).
У губок є декілька варіантів протікання метаморфозу. У більшості вапнякових губок при повному рівномірному дробленні утворюється бластула з однакових джгутикових клітин, яка починає вільно плавати. Далі частина клітин втрачає джгутики й іммігрує всередину бластули. Порожнина бластули повністю заповнюється амебоїдними клітинами. Утворюється паренхімула, яка через деякий час осідає на дно і перетворюється на маленьку губку. У звичайних, скляних і деяких вапнякових губок при повному нерівномірному дробленні утворюється амфібластула з дрібними джгутиковими клітинами (мікромерами) на одному кінці бластули й великими безджгутиковими клітинами (макромерами) на другому кінці. Прикріпившись мікромерами до субстрату, вона перетворюється на молоду губку. У багатьох невапнякових губок з яйця виходит личинка паренхімула, в якої дуже швидко відбувається раннє диференціювання клітин.
У кишковопорожнинних у класу гідроїдних поліпів з яйця виходить паренхімула, подібна до личинки губок. Вона має поверхневий шар клітин з джгутиками та внутрішній шар клітин. Пізніше у внутрішньому шарі клітин утворюється гастральна порожнина; утворюється планула. Планула через деякий час прикріплюється переднім кінцем. На протилежному кінці проривається рот, навколо нього виростають щупальця; утворюється молодий поліп. У сцифомедуз та коралових поліпів з яйця виходить одразу планула. У деяких коралових поліпів виходить ще більш пізня личинка, яка має щупальця і нагадує поліпа; вона називається актинулою.
У голкошкірих постембріональний розвиток розпочинається зі стадії бластулоподібної личинки, клітини якої мають джгутики. Далі шляхом імміграції та інвагінації відбувається гаструляція. Первинний кишечник розділяється на зачаток целомічної мезодерми і зачаток кишечника. Шляхом імміграції утворюється мезенхіма. Бластопор перетворюється на анальний отвір, а на протилежному кінці утворюється рот. Личинка починає самостійно харчуватися. Потім утворюється навколоротова впадина, по краю якої проходить кайма особливо довгих джгутиків – війчастий шнур. Утворюється диплеврула. Потім утворюються вирости та лопасті, на які переходить і війчастий шнур.
Така личинка має трохи різну будову у різних голкошкірих і називається по-різному: біпінарія (у морських зірок), аурикулярія (у голотурій), плутеус (у морських їжаків та офіур), бочковидна личинка (у морських лілій). Всі ці личинки двобічносиметричні. Целомічний мішок розділяється на три пари целомів. Як правило, краще розвинуті целоми лівого боку. Розпочинається власне метаморфоз. Середній лівий целом (гідроцель) оточує стравохід і замикається в кільце (утворюється навколоротове кільце амбулакральної системи). Від нього відростає 5 вип’ячувань – зачатки радіальних каналів, навколо яких формуються амбулакральні ніжки. Радіальну симетрію стають мати кровоносна й нервова системи. Метаморфоз завершується і утворюється молода морська зірка.
У плоских червів, що ведуть паразитичний спосіб життя, складний життєвий цикл, в якому, як правило, зустічається личинкова стадія і відбувається метаморфоз. У більшості ж вільноживучих плоских червів розвиток прямий.
У кільчастих червів з метаморфозом розвиваються багатощетинкові (Polychaeta). При цьому з яйця виходить личинка трохофора (мал. 1а). Це типова планктонна личинка, що рухається за допомогою війок. Тіло її більш-менш кулеподібне або дещо витягнуте, на верхньому полюсі розташована тім’яна китиця війок, які сидять на групі ектодермальних клітин. Це орган чуття, біля нього часто лежать інші органи чуття: пара вічок, коротенькі щупальця, статоцисти. Приблизно по екватору трохофори проходить кільце війок – прототрох, або передротовий віночок, який поділяє тіло личинки на верхню півкулю – епісферу та нижню – гіпосферу. Безпосередньо під прототрохом розташований рот, на нижньому (вегетативному) полюсі – анус.
Крім прототроха, у багатьох трохофор є ще додаткові війчасті кільця: метатрох, або післяротовий віночок, що міститься позаду рота, та телотрох – перед анальним отвором. В епісфері, під тім’яною пластинкою, лежить нервовий ганглій, що іннервує органи чуття личинки. З ним зв’язані нервові меридіональні і кільцеві стовбури. Кишечник трохофори має вигляд вигнутої трубки і складається з ектодермального стравоходу, ентодермальїюї середньої кишки та ектодермальної задньої. Між стінкою тіла трохофори і кишечником розташована первинна порожнина тіла, в якій містяться поодинокі мезодермальні клітини (мезенхіма) та м’язи. У нижній півкулі личинки є пара протонефридіїв. Біля нижнього полюса трохофори з обох боків від кишечника лежать дві великі клітини – мезодермальні телобласти, іноді вони починають ділитися, утворюючи дві мезодермальні смужки.
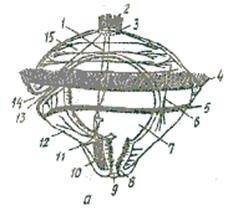 | 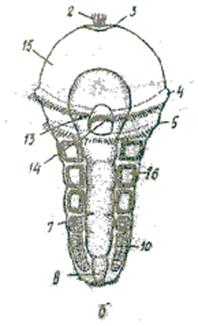 | ||
Мал. 1. Личинки у кільчастих червів, ракоподібних та молюсків: А – трохофора; В – метатрохофора (наупліус). 1 – нервові стовбури; 2 – тім’яна китиця; 3 – тім’яна пластинка; 4 – прототрох; 5 – метатрох; 6 – м’язи; 7 – середня кишка; 8 – задня кишка; 9 – анус; 10 – мезодермальна смужка; 11 – протонефридій; 12 – гіпосфера; 13 – передня кишка; 14 – рот; 15 – епісфера; 16 – целомічні мішки.
Після деякого періоду планктонного життя починається метаморфоз трохофори. Задня половина тіла личинки значно подовжується і поділяється одночасно на кілька (найчастіше 3, 7 або 13) сегментів, на яких розвиваються зачатки параподій та щетинки або віночки війок. У цей час телобласти починають ділитись, утворюючи два тяжі – мезодермальні смужки, що лежать по боках від кишечника. Якщо ці смужки вже були у трохофори, то тепер їх клітини посилено діляться. Згодом усередині кожної з мезодермальних смужок клітини розсуваються, й утворюється порожнина – зачаток целома.
Найчастіше целомічні мішки залишаються у личинки суцільними, проте в деяких видів вони розділяються, утворюючи по парі целомів у кожному сегменті. Личинку, що має кілька сегментів, називають метатрохофорою (мал. 1б). Її тіло складається з головної лопаті (простомія), яка становить майже незмінену верхню півкулю трохофори, кількох сегментів і маленької анальної лопаті (пігідія) – ділянки нижньої півкулі трохофори навколо ануса. Сегменти, що угворились у метатрохофори, називають личинковими, або ларвальними. У багатьох видів поліхет метатрохофора перетворюється на наступну личинкову стадію – нектохету, в якої вже з’являються головні придатки – антени, пальпи, лопаті параподій із щетинками, тобто вона більше схожа на дорослу особину, але має лише ларвальні сегменти і веде планктонний спосіб життя.
На епісфері в неї розвиваються очі, антени, органи нюху. Зв’язані з ними нервові клітини, об’єднуючись, утворюють головний мозок (надглотковий ганглій). В ектодермі черевної сторони тіла з’являються зачатки гангліїв черевного нервового ланцюжка. Згодом на задньому кінці личинки, перед анальним отвором утворюється зона росту, спереду від якої один за одним формуються сегменти, що звуться постларвальними. У зоні росту клітини мезодермальних смужок інтенсивно діляться, від них відділяються парні зачатки целомічних мішків. Кожен сегмент, що утворюється, має пару таких зачатків, які, розростаючись, утворюють целомічні мішки.
Зовнішньою стороною кожен мішок підстилає шкірно-м’язовий мішок відповідного сегмента, внутрішньою оточує кишечник. Стінками целомічних мішків, що прилягають один до одного над і під кишечником, утворюються спинна і черевна брижі, а в місці стикання розташованих один за одним мішків – поперечні перетинки між сегментами – септи, або дисепименти. Кровоносні судини виникають між двома шарами целомічного епітелію у спинній та черевній брижах (мезентеріях). За рахунок клітин мезодермальних смужок формуються також мускулатура шкірно-м’язового мішка і кишечника, клітини целомічної рідини, крові, хлорагогенні клітини. Утворення сегментів у зоні росту триває дуже довго, іноді до кінця життя черва, що призводить до значного збільшення їх кількості.
У підтипу ракоподібних, який відноситься до типу членистоногих, більшість представників розвивається з метаморфозом. У різних груп він відбувається по-різному. Перша личинка, що виходить з яйця, називається наупліусом. Вона овальної форми і має три пари кінцівок – антенули, антени та мандибули (верхні щелепи), останні дві пари двогіллясті. На відміну від дорослих форм, у наупліуса антени лежать на черевній стороні тіла, по боках від рота, і призначені для плавання та збирання іжі, а мандибули спочатку – тільки для плавання.
На задньому кінці тіла личинки є анальна лопать (тельсон). Наупліус за будовою відповідає метатрохофорі поліхет (мал. 2б). Наупліалькі сегменти вважаються ларвальними, а всі інші, що виникають пізніше,– постларвальними. Щодо кількості ларвальних сегментів у ракоподібних єдиної думки немає. Науптіус має кишечник, головний мозок, два черевні ганглії, непарне наупліальне око й одну пару видільних органів, найчастіше це антенальні залози. На задньому кінці тіла, між мандибулярним сегментом і тельсоном, розташована зона росту. Тут утворюються наступні сегменти тіла. Наупліуси ведуть планктонне життя й розносяться (особливо морськими течіями) на далеку відстань.
Наупліус росте, в зоні росту утворюються нові сегменти із зачатками кінцівок: спочатку сегменти з першою і другою парою максил (нижні щелепи), а потім – передні грудні. Личинка, в якої починають формуватися постларвальні сегмента, зветься метанаупліусом. Під час постембріонального розвитку відбуваються линяння. Після кожного з них з’являються нові сегменти і закладаються нові кінцівки, тобто розвиток відбувається шляхом анаморфозу. При цьому можуть змінюватися будова та функції кінцівок. Наприклад, усі три пари кінцівок наупліуса, як уже згадувалося, призначені для плавання; пізніше ж антенули і антени стають органами чуття, а мандибули – ротовими придатками. Поступово кінцівки набувають остаточного вигляду, і метаморфоз закінчується.
Постембріональний розвиток у класі Malacostraca (Вищі раки) має свої особливості. У них скорочується й стабілізується кількість линянь, утворюються додаткові личинкові стадії. Часто спостерігається ембріонізація розвитку, в результаті якої ранні личинкові стадії (наупліус та метанаупліус) проходять у яйці, а назовні виходить більш пізня личинка (найчастіше, зоеа). Зоеа – це личинка, що вже має всі сегменти, але середні сегменти ще повністю не сформовані й не мають кінцівок. Зоеа має розвинені ротові кінцівки й ногощелепи, а також зачатки грудних ніжок і сформоване черевце, на якому є лише остання пара кінцівок.
На передній частині тіла є фасеткові очі. Зоеа линяє й переходить у так звану мізидну стадію: у неї вже є повністю сформовані двогіллясті грудні ноги (як в представників ряду Mysidacea, звідки й назва личинки) й зачатки черевних кінцівок. У різних представників десятиногих раків мізидна стадія має різну будову і відповідно різні назви: у крабів – мегалопа, що зовні нагадує дорослу тварину, але має витягнуте черевце; у лангустів – філосома, яка має прозоре листкоподібне тіло, довгі двогіллясті грудні кінцівки й дуже коротке почленоване черевце; тощо.
Мізидна стадія після линяння перетворюється на цілком сформовану тварину. Деякі вищі раки, наприклад, креветки з родинм Penaeidae, проходять усі личинкові стадії: наупліус, метанаупліус та наступну мізидну стадію. У ряді груп метаморфоз відбувається дуже своєрідно. У більшості видів ряду Decapoda (десятиногі раки), а також у деяких інших таксонах спостерігається повна ембріонізація розвитку, в результаті чого з яйця виходить мініатюрна копія дорослої тварини з усіма сегментами й сформованими кінцівками.
Комахи
Серед безхребетних непрямий розвиток найкраще вивчений у комах. У них виділяють два основних типи метаморфозу: 1) з повним перетворенням; 2) з неповним перетворенням.
При неповному перетворенні (геміметаболії) комаха проходить такі фази розвитку: яйце, личинка, імаго. З яйця виходить личинка, зовні доволі схожа на дорослу особину. Вона має фасеткові очі, такий самий як і в дорослих ротовий апарат, а на старших стадіях – зовнішні зачатки крил. Спосіб життя личинок багатьох видів з неповним перетворенням і дорослих комах однаковий. Ці личинки звуться німфами. Окрему модифікацію становлять так звані наяди – личинки одноденок, бабок і веснянок.
Вони теж схожі на імаго мають фасеткові очі й на старших стадіях – зачатки крил, але живуть у воді й мають спеціальні личинкові органи – зябра, маску (личинки бабок) тощо. Ріст і розвиток личинки супроводжуються періодичними линяннями, кількість яких, як правило, стала для певних таксонів (найчастіше, родин). З кожним линянням личинка за розмірами, пропорціями тіла, розвитком крил наближається до дорослої фази; під час останнього линяння вона перетворюється на імаго.
При повному перетворенні (голометаболії) комахи проходять такі фази: яйце, личинка, лялечка, імаго. З яйця виходить личинка, яка різко відрізняється від імаго більшою гомономністю метамерії, відсутністю зовнішніх зачатків крил, слабкішим розвитком органів чуття. Вона не має фасеткових очей, антени її значно коротші; часто ротовий апарат у неї інший, ніж у дорослої комахи (наприклад, у гусені метеликів – гризучий, а в імаго – сисний). Личинки комах із повним перетворенням живуть в інших умовах, ніж дорослі, і з ними не зустрічаються. Більшість органів личинки має тимчасовий характер, наприклад черевні ноги в личинок метеликів та пильщиків, шовковидільні залози у гусені метеликів. Личинкові ознаки зберігаються під час усієї личинкової фази.
Із кожним линянням личинка лише збільшується в розмірах, а необхідні для метаморфозу зміни відсуваються до останнього личинкового линяння. Закінчивши свій ріст, личинка останньої стадії припиняє живлення, стає нерухомою, линяє востаннє й перетворюється на лялечку. Лялечка нерухома, але це тільки зовнішнє враження: на цій фазі відбуваються інтенсивні процеси внутрішньої перебудови – гістоліз та гістогенез. Оскільки характер руху імаго (особливо, політ) і його живлення інакші, ніж у личинкової фази, то м’язи, травна система, ротовий апарат, кінцівки та інші личинкові органи мають зовсім перебудуватися. Більшість органів личинки руйнується; цей процес зветься гістолізом.
Він відбувається шляхом фагоцитозу, аутолізу (самоперетравлення), а також,– за допомогою ферментів. У різних комах переважає той чи інший механізм, але найчастіше діють усі три: гемоцити виділяють у гемолімфу ферменти, які руйнують тканини; їх залишки фагоцитуються гемоцитами. Крім того, в деяких тканинах відбувається аутоліз. Коли гістоліз досягає свого піку, внутрішні органи лялечки перетворюються на напіврідку масу, яка складається з гемолімфи, збагаченої продуктами розпаду. Не руйнуються лише нервова, статева системи та спинна кровоносна судина. Нервова система може доповнюватися новими клітинами, в ній можуть концентруватися ганглії, проте вона ніколи не втрачає своєї цілісності. Паралельно з гістолізом відбувається гістогенез – побудова імагінальних органів.
Органи дорослої комахи формуються з особливих зачатків – імагінальних дисків. Це невеличкі скупчення недиференційованих клітин, розташовані в певних місцях тіла личинки. Кожен імагінальний диск має своє призначення: є диски, з яких утворюються крила, кінцівки, певні ділянки кишечника, фасеткові очі тощо. Імагінальні диски закладаються ще під час ембріонального розвитку або в ранніх личинок; протягом личинкового розвитку вони ростуть, але не диференціюються. Тільки в личинки останньої стадії перед залялькуванням їхні клітини диференціюються; в лялечки вони розгортаються, утворюючи імагінальні органи. Важливу роль у метаморфозі відіграє жирове тіло. На личинковій фазі в ньому нагромаджуються поживні речовини (жири, білки, вуглеводи), що використовуються як пластичний матеріал і джерело енергії при побудові імагінальних органів.
Метаморфоз комах відбувається під гормональним контролем з боку нейроендокринної системи (мал. 2). Метаморфоз регулюється нейроендокринним комплексом органів, до якого входять нейросекреторні клітини мозку, кардіальні тіла, прилеглі тіла та парна проторакальна (передньогрудна) залоза. Нейросекреторні клітини мозку виробляють проторакотропний гормон. Через довгі аксони цих клітин він надходить до кардіальних тіл, а з них – у гемолімфу. У личинок проторакотропний гормон стимулює роботу проторакальних залоз, які починають виділяти гормон линьки – екдізон. Прилеглі тіла виділяють ювенільний гормон.
Концентрація екдізону мала в періоди між линьками і висока – в період линьки; а концентрація ювенільного гормона висока протягом всієї личинкової стадії, і знижується перед линянням на імаго чи лялечку. Екдізон в периферичних тканинах перетворюється на свою активну форму – екдістерон і впливає на клітини гіподерми, спричиняючи спочатку синтез ферментів, які розчиняють стару кутикулу, а пізніше – синтез матеріалу для побудови нової кутикули. Крім того, екдізон стимулює ріст і диференціацію всіх тканин личинки, в тому числі,– й клітин імагінальних дисків.
Але при високій концентрації ювенільного гормону екдізон не може активувати розвиток імагінальних дисків і тому стимулює лише секрецію личинкової кутикули, і линяння завершується появою чергової личинкової стадії. Зниження концентрації ювенільного гормону веде до зняття блокуючого впливу на імагінальні диски, екдізон стимулює їх ріст та диференціацію, і відбувається линяння личинки на лялечку (або останню личинкову стадію при неповному метаморфозі).