 2015-06-28
2015-06-28 4678
4678Цитоплазма түссіз сұйьщ гиалоплазма мен органоидтардан тұрады. Клетканьң өзіне
тән органоид — мембраналар системасы, олар клетка мембранасыньң жалғасы болып табылады, олардың үсті тегіс болады немесе үстінде диаметрі 10—15 нм ете ұсақ гранулалар (рибосомалар) болғандықтан, бұдыр-бұдыр болып келеді Осы мембраналар системасы эндоплазмалық тор деп аталады. Ол тор клеткалық және ядролық мембраналарды байланыстырады және оның табиғаты сол мембраналардыы сияқты липопротеин тектес болады. Эндоплазмалық, тормен — мембраналардан түзілген түтіктер арқылы клетка ішіндегі заттар қозғалады. Бұдан өзге мембраналар системасының маңызды ерекшелігі клеткаларды тұйық, кеністігі бар бөліктерге бөліп тұрады.Соңғы жылдары электрондық микроскоптың көмегімен клеткаларда цитоплазмалқщ жіпшелер мен микротүтікшелер табылды. Клеткаларда жіпшелердің рөлі ерекше зор: мысалы, олар бұлшық ет талшықтарындары жиырылу механизмінің компоненті болып табылады. Олар клетканьң қатаңдығын сақтайды және жыртылуға төзімді етеді.Белгілі 6ip бағыты бар түтікті құрылымды микротупкшелер деп атайды. Олар эукариоттар клеткаларының универсал компонентін құрайды. Түтіктер диаметрі 20—27 нм. Олардын қабырғаларының қальңдығы 5—7 им шамасындай, 6ip 10 және одан да көбірек жіпшелерден тұрады. Микротүі!кшелер мен цитоплазмалық жіпшелердің байланысы әлі жете зерттелмеген. Олардың клеткадғгы рөлө алуан түрлі. Ен алдымен олар жіпшелермен қосылып цитоскелет құрайды, клетка қозғалысы (мысалы, кірпікшелер құрамына кіреді) және клетка бөлінуі кезіндеті хромосомалар қозғалысы механизмінің бөлшектері (бөқліну ұршығының құрамына кіреді) болып табылады.Цитоплазмада өзінің мембранасына қоршалған басқа органоидтар: митохондриялар, рибосомалар.Гольджи аппараты, лизосомалар, жануарлар мен төмеңгі сатыдағы өсімдік клеткаларындағы центросома, өсімдіктерден пластидтер және т. б. болады. Митохондриялардың формалары таяқша тәріздес және гранула пішіндес болады. Олардың кеністікпен бөлінген екі мебранадан тұратын қабьқшасы болады. Сыртқы мембрана 6ip тепе жылтыр, ішкi мембрананың, көптеген катпарлары — кристалдары болады. Әр организм түрше және тканьдер типінепішіні,
үлкенді әр түрлі және биохимиялық, процестерден митохондриялар рөлі тән болады.
Клетканың функционалдық, жағдайына байланысты олардын, формасы да өзгеріп отырады. Митохондриялардьң үлкенді шамасы едәуір денгейде: 0,2-ден 2— 7 мкм шамасына дейш ауытқып отырады. Клеткадағы тотығу арқылы зат алмасуға олардын қатысатыны анықталды.
Митохондрияда кребс циклінің барлық, реакциялары жүреді деген болжам бар. Митохондриялар — энергия дайындаушылар. Рибосомалар цитоплазмада (негізінде мембрананын, үстінде) сондай-ақ, олар ядрода, пластидтерде және митохондрияларда едәуір мөлшерде кездесеі1. Олар мөлшері әр түрлі ей суббірліктен тұрады. Жалпы мөлшері 15-тен 35 нм аралығында, жарьқ микроскоптан рибосомалар көрінбейді Олардын ерекшелігі көп мөлшерде РНК (рибосомалық,) және белоктар болады; бүкіл клеткалық РНК 80—90% рибосомаларда болады. Рибосомалар, ядроньң бақылауымен клетка белоктарының, синтезше қатысатыны аньқталды. Гольджи аппараты клетка мембранасы системасымен байланысты және клетканың, бөліп шығару және секреция функциясын қамтамасыз етеді Лизосомалар — біқатар ферменттерден тұратын және өздеpi клетка ішіндегі ас қорыту функциясын атқаратын дене: ол ферменттер үлкен молекулалардың ұсақ белштерге ажырап бөлінуіне себеп болады, олар митохондрия ферменттерімен тотығады. Лизосоманы қоршаған мембрананың жыртылуы клеткалар лизисына әкелiп соғады. Центросома (клетка орталығы) мынадай екі компоненттен: шағын денелер — центриолдардан және центросфералардан (цитоплазманың ерекше жағдайда жіктелген бөлігі) тұрады. Клетканың, бөлінуі кезеңінде пайда болатын ахроматин ұршығының қалыптасуы центросомға байланысты. Гүлді өсімдіктер клеткасынан центросомалар табылған жоқ,: мұнда ахроматин ұршғгы клетка полюстерінде «поляр қалпақшалары» түрінде пайда болады. Пластидтер (хлоропластар, лейкопласта р, хромопластар т. с. с.) өсімдік клеткалары цитоплазмасына тән, олардың, құрылысы ете күрделі. Олар фотосинтезі1, крахмал мен пигменттер синтезін, сонымен қатар липидтер, пластидті нуклеин қышқылдары және белоктар синтезін іске асырады. Клетка мембраиасының құрылысы күрделі болады, олардың құрылысы қоғганыш, талғап өткізу және бөлшектер мен молекулаларды активті тартып, өзіне енгізу сияқты белгілі 6ip функция атқаруға бейімделген.Жалпы алғанда, клетканы тіршіліктің ен, қарапайым бөлігі деуге болады: өйткені клеткада зат алмасуға және көбеюге қажеттінің 6әpi де бар.
Клетканың, жалпы құрылымың қысқаша еске түcipy, ары қарай клетканың жеке
элементтері тұқым қуалаудағы рөлі бағалау үлкен қажет болды.
Клетка цикліндегі оның, құрылымыньң динамикасы және хромосома құрылысы келесі тарауда қарастырылады.
2-тарау. ЖЫНЫССЫЗ КӨБЕЮДІҢ
ЦИТОЛОГИЯЛЫҚ НЕГІЗДЕРІ
Организмнің, жыныссыз және вегетативтік көбеюінің негізше универсал процесс — клетканың, бөлінуі жатады.
I. КЛЕТКАНЫҢ БӨЛ1НУІ
Митоз. Клетканың, бөлінуі —• организм көбеюінің орталық, кезені. Клетканың, бөлінуі нәтижесінде 6ip клеткадан екі клетка пайда болады.
Бұл процесс негізгі екі кезеңен тұрады: ядроньң бөлні — митоз (кариокинез) деп, цитоплазманың,бөлінуі— цитокинез. Клетка өзінің тіршілік циклінде кезектесіп келетін алты стадияны басынан өткізеді: интерфаза, профаза, прометафаза, метафаза, анафаза және телефаза (I таблица, А). Бұл стадиялардың 6әpi қосылып интерфаза мен митозра жіктелетін 6ip митозды, цикл құрайды.Клетканың екіге бөлінуі арасында ядро интерфаза стадиясында болады. Интерфазада клетканың, ерекшелігіне тән және клетканың бөлінуіне қажетті заттар түзшеді. Бұл кезде ядродан ұсақ, жіпшелерден — хромосомалардан құралған тор құрылымы жақсы көрінеді. Профазада — митоздьқ 6іріншi кезеңінде хромосомолар спиральданады да, екіден қосарланган жіп сияқты болып жарық, микроскопынан көрінеді.Интерфаза кезінде хромосоманың қосарлануы немесе оның репродукциясы болатынын байқаймыз. Бұл кезде бастапқы хромосомалардын, әркайсысы дәл өзі сиқты жаңа хромосома түзеді: Сіңлілі хроматидтер деп аталатын бұл жарты бөлік профаза кезінде бөлініп кетпейді, оларды центромера (кинетохором) деп аталатын ортақ бөлік біріктіріпұcтап тұрады.Профазада хромосомалар ары қарай ұзынынан спиральдана түседі, соньң нәтижесінде олар қысқарады және жуандайды. Сол сияқлы профазада хромосомалар ядроның бүкіл келемше таралатынын атап көрсету маңызды.Жануарлар клеткасында интерфазанын, бас кезінде немесе тіпті телефазалық, бөлінудің, кезінде центриолдар қосарланады, бұдан кейін профазада жас центриолдар ажырап, клетканьң полюсіне қарай бөліне бастайды. Центриолдар арасында бір буда бөліну ұршығының жіңішке жіпшелері пайда болады, осы жиынтық ахроматин аппараты деп аталады.
Бұлшық ет клеткалары құрамындағыдай, ұршық жіптері құрамында актин белогы
болады, ол белок қозғалыстың түрлі жағдайында жиырылуды қамтамасыз етеді.
Профазаның аяқталуының негізгі белгісі — ядрошықтар мен ядро қабықшасы жоғалып кетеді, сонын, нәтижесінде хромосо-малар цитоплазма мен нуклеоплазманың жалпы массасының ішінде орналасады.
Прометафаза клеткадағы хромосомалардың экватор жазықтығына қарай қозғалуымен сипатталады. Бұл қозғалыс пен хромосомалардың экватор ұршығында таралуы метакинез деп аталады.ОЧетафаза деп хромосомалардың экватор жазықтығында ұршық өсіне перпендикуляр орналасуын айтады. Осы жазықтыққа орналасқан хромосомалар экваторлық немесе метафазалық пластинка құрайды. Әрбір хромосома экваторлық жазықтыққа оның центрлері дәл келетіндей болып орналасады, ал хромосомалардың қалған барлық денесі одан тыс жатуы мүмкін. Экваторлық пластинканы клетканың бөлінуі полюсінен қараған кезде барлық хромосомалар жақсы көрінеді, оларды санауға және формасын байқап көруге болады. Цитоплазманың қалған массасына қарағанда ұршық жіптері тығыз консистенциялана түседі. Олар хромосомаларға мынадай жолмен, яғни центромераға "жіп екі полюстен келіп жалғасады.Митоздың келесі фазасы анафаза деп аталады, бүл кезде центромералар және сіңлілі хроматидтер (оларды енді хромосомалар деп атауға болады) бөлінеді де, полюстерге таралады. Мұнда ең алдымен хромосоманың центромералық учаскелері бірінен-бірі алшақтайды, б9дан кейін алдымен центромерлер, сонан соң хромосомалардың өздері полюстерге ажырайды. Анафазада хромосоманың ажырап бөлінуі — «комаыда берілгендей» — бір мезгілде басталады да, өте тез аяқталады. Хромосомалар ажырап барғаннан кейін, екі полюстегі олардың саны бірдей болады және әр бөліктегі хромосом саны бастапкы клеткадағы хромосома санына тең болады. Ядро бәлінуінің осығідай ерекшелігіне байланысты клетка ұрпақтарында хромосома саны және олардың сапалық құрамы үнемі түрақты болады.
Телефазада жас хромосомалар деспиральданады. дараланып көрінуі жойылады. Ядро қабықшасы пайда болады. Бұдан кейін ядрошық (немесе ядрошықтар) қалпына келеді, оның саны бастапқы ядродағыдай болады. Ядро енді профазада болған езге-ріспен салыстырғанда кері реконструкцияланады.
Цитокинез. Пластидтер бөліну арқылы көбейеді, сірә олар клеткада жаңадан пайда болмаса керек. Митохондриялар бөліну арқылы көбейеді деген болжам бар. Жалпы алғанда клетка органоидтары репродукциясының механизмі жөнінде мәліметтер өте аз. Бұл процесс асинхронды жүреді, ал ол бөліктердің бөліну жылдамдығы ядро
бөлінуінің жылдамдығымен дәл келмейді деген мәлімет бар.
Цитокинез кезінде органоидтардың жа-ңа пайда болған жас клеткаларға бөлінуінде қатал заңдылық жоқ, сірә бұл процесті бак,ылайтын арнайы механизм болмаса керек. Осыған байланысты жас клеткалардағы бір аттас органоидтардың саны бірдей болмайды. Органоидтардың жас клеткалар арасында теңдей бөлінбеуі, олардың кездейсоқ таралуы клеткалардың тіршілігін бұза алмайды. Сірә, клеткада бірін-бірі алмастыратын аттас бөлшектер өте кеп болады.
Клетка денесінің бәлінуі — цитокинез — ядро бөлінуінен кейін ілешала басталады. Жануарлар клеткасының бөлінуі аналық клетканың экваторы бойынша цитоплазманың шетінен ортасына қарай буыақталу жолымен жүреді.
Өсімдік клеткасында клеткалық аралықтың қалыптасуына ортасынан шетіне бунақталатын фрагмопласт деп аталатын ұршықтың қатысымен жүреді. Осымен митоз аяқталады.Митоздық циклдің ұзақтығы организм түріне, ткағіь типіне, организмнің физиологиялық күйіне, сыртқы факторларға (температураға, жарық режиміне т. с. с.) байланысты бірнеше минуттан бірнеше тәулік шамасында ауытқиды. Митоздың жеке фазаларының ету жылдамдығы өзгергіш болады.Клетканың бөлінуге дайындығын анықтайтын себептер, оиы жүргізуші механизм осы уақытқа дейін анықталған жоқ.
Қлетка бөлінуінің митоздан басқа типтері де белгілі. Амитоз. Ахроматин үршығын түзбей, ядроның тікелей бәлінуі амитоз деп аталады. Бөліну ядроның бунақталып екі бөлікке бөлінуімен жүреді, кейде бір ядродан бірден бірнеше ядро түзіледі (фрагментация). Клетканың амитоз жолымен бәлінуі бірқатар арнайы клеткаларда және патологиялық тканьдерде үнемі кездеседі. Мысалы, картоптың крахмал түзуші клеткаларында, регенерация кезіндегі бұлшық ет клеткаларында, рак клеткаларында, қарапайым организмдерде болады.
Эндомитоз. Эндомитоз деп клеткадағы хромосоманың репро-дукциялануы кйзінде ядроның бәлінбей жүру процесін айтады. Осының нәтижерінде клеткада хромосома саны көбейеді, кейде ол бастапқы санымен салыстырғанда ондаған есе артады. Эндомитоз өсімдіктердегі сияқты жануарлардың да әр түрлі тканьдердегі белсенді жұмыс істейтін клеткаларында кездеседі. Кейде хромосомалардың пайда болуы клеткадағы олардың санының өсуінсіз өтеді. Бұл кезде әр хромосома еселеп еседі, бірақ жас хромосома өзара байланысқан күйінде қалады. Бұл құбылыс лолитения деп аталады. Ол эндомитоздың жеке жағдайынан тұрады. Политенді хромосомада жіпшелер саны 1000—2000 шамасына жетеді. Бұл кезде аса зор алып хромосомалар түзіледі.
Политения құбылысы бірқатар диффереицияланған тканьлеткаларда байқалады және ол клетка ядросының ерекше байланысты болады.
Клетканың белінуі процесіндегі болатын ерекше ды бағалау үшін,хромосоманың құрылысын және ұдайы дамып отыруын яғни оның репродукциясын жан-жақты қарастыру қажет.
2. ХРОМОСОМАЛАРДЫҢ ҚҰРЫЛЫСЫ ЖӘНЕ ОЛАРДЫҢ РЕПРОДУКЦИЯСЫ
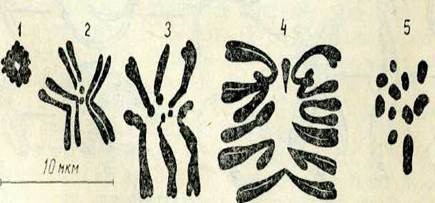
Хромосомалардың морфологиясы және көлемі. Хромосомалардың жалпы морфологиясы оларды ең қысқарған және экватор жазықтығында орналасқан метафаза немесе анафазаның бастапқы стадиясында жақсы байқалады. Бұл кезде хромосомалар жақсы көрінеді, олар пішіні мен үлкендігі жөнінен айрықшаланады (2-сурет). Әр хромосоманың формасы негізінен алғашқы тартылыс кезінде анықталады, ол жерде центромера бо-лады. Әр түрлі хромосомолардағы центромераның орны түрліше болады, бірақ ол әр хромосоманың өзіне тән және тұрақты болады.
Центромера хромосоманың ең ұшында болмайды деп бол-жайдьц Хромосомалардың ең үшындағы сеғменттері теломера ' деп
аталады./Хромосоманың центромераға жақын орналасқан. учаскесін проксимальді, қашығырақ учаскесін дистальды деп атайды.
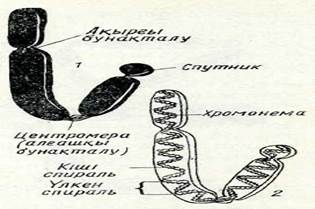
Хромосомада алғашқы тартылыстан өзге, ұршық жіпшелерімен байланыссыз екінші тартылыс болуы мүмкін, (2-сурет, 5). Хромосомадағы бұл тартылыс ядрошықтың қалыптасуымен байланыстьу оны ядрошық уйымдастырушы деп атайды.;Хромосоманың бұл учаскесінің құрылысы күрделі екені анықталды жәнерибосомалық РНК синтезіне жауапты.ІЕкінші тартылыс кейде өте ұзын болуы мүмкін, ондай жағдайда ол тартылыс хромосоманың негізгі денесінен шағын учаскені бөліп тұрады, ол спутник деп аталады Мұндай хромосомалар спутникті деп аталады / (2-сурет, 6).
Жоғарыда айтылғандай, метафаза кезінде хромосомаларды полюске тарататын центромераға ұршық жіпшелері бекітіледі. Егер хромосоманың центромерсіз учаскесі (ацентрлік фрагмент) бүзылса, ол репродукциялана алуға қабілетті, бірақ центромераны қалпына келтіре алмайды. Центромерасы болмағандықтан, мұндай ацентрлік фрагмент ориентация жасай алмайды, сондықтан клетка бөлінуі кезінде көбінесе жойылып кетеді.
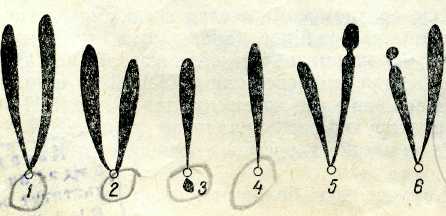 |
2-сурет. Метафазалық хромосомалар типі:
1 — метацентрлік (екі бөлігі бірдей); 2 —субметацентрлік; 3— акроцентрлік (екі белігі мүлде бірдей емес); 4 — телоцентрлік (екінші бөлігі;қоқ) хромосома; 5 — екінщі тартылыстіл дентрлік хромосома; б— спутникті хромосома. Ашық түсті дөңгелектер арұқылы центромера көрсетілген.
 |
Фрагментцентромерасымен басқа хромосомаға байланысса ғана сақталады.
Алайда таякша тәріздес хромосомалар да белгілі, ұршық жіпшелері оның бүкіл денесінің ұзына бойына жалғасады. Мұндай хромосомалардың диффузды центромерасы болады. Мұндай жағдайда бөлек қалған хромосома фрагменттері анафазаға бір қалыпты ажырауы мүмкін. Бүл құбылыстың та-биғаты әлі жете зерттелмеген. Хромосомалардын, морфологиясы ғана емес, үлкендігі де әр түрлі. Олардың ұзындығы 0,2—50 мкм аралығында; диаметрі 0,2—5 мкм (немесе 20—50 нм). Алайда белгілі бір хромосоманын, ұзындығы біршама түрақты.
Жіктеп бояуға мүмкіндік беретін хромосоманы бояудың жаңа әдістерін
қолдану, яғни хромосоманың боялған және боялмаған бөліктерінің заңды кезектесуін — диаметрі әр түрлі дискілерін анықтау, әр хромосомаға дәл сипаттама беруге яғни хромосоманың жеке-даралығын анықтауға мүмкіндік берді (3-сурет).
Сонымен әр клеткада хромосомалардың формасын, үлкендігін және жіктеліп боялуын қарастырған кезде олардың бәрін өзара ұқсатуға болады. Ал зерттеп білуді жеңілдету үшін, әр-қайсысын ажыратып нөмірлеу керек. Мысалы, адам хромосома-лары (28-тарауды қараңыз) және бірқатар басқа организм хромосовдалары осылай істелген.
Хромосомалардың құрылымы. Бөлінетін клеткалардағы хро-мосомалардың құрылымы профазада көріне бастайды. Жоғарыда айтылғандай, алғашқы профазада хромосомалар қосарланған жіңішке жіп (сіңлілі хроматидтер.) тәріздес болады. Метафаза стадиясында хромосомалардың 4 жіпшеден тұратыны кейде жарық микроскопынан кәрінеді, олар жартыхроматидтер деп аталады. Жартыхроматидтер хромосома қүрылымындағы ең қарапайым ұсақ бөлік емес екені электрондық микроскоп кемегімен анықталды. Әр хромосоманың өзі хромонема деп аталатын кептеген жіпшелерден құралады деген гипотеза айтылып жүр. Рентгеноструктуралық анализ мәліметтері бойынша ең жіңішке жіпшенің диаметрі 3 ым шамасындай болады. Хромосоманың кептеген жіпшелерден қүұралған құрылымы 4-суреттен жақсы көрінеді. Осы модельге сәйкес хромосомадағы жіпшелердің жалпы саны 64.
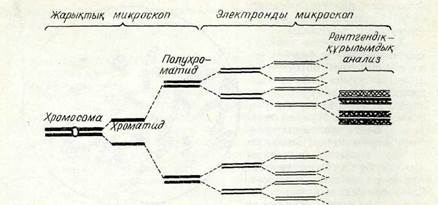
Бір жіпшенің дисшетрі НМ: 20 -25 НМ 10 -12 НМ 3 -4-НМ
ДНКмошулалары-нық. саны
| 6-сурет. Метафазалық хромосома |
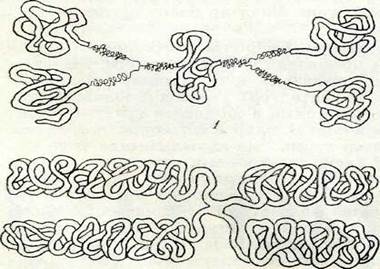 |
4-сурет. Хромосоманьщ мпкроскоптық, субмикроскоптык, және молекулалық қурылымының схемасы (көп жіпшелі моделі) Сол сияқты екінші гипотеза да белгілІ, ол бойынша хромо-сомалар бір ғана жіптен тұрады, сол жіптің спиральдануы және бүктелуі нәтижесінде политенді құрылым қалыптасады (5-су-рет). Әр түрлі организмдер клеткасының ядросында ұзындығы 1—22 мм жететін ДНҚ молекуласының үздіксіздігі осы соңғы гипотезаны қоштайды.
 |
5-сурет. Хромосоманың бір жиішелі құрылым схемасы (қабаттасқан жіпше);
1 — репродукция ироцесі кезіндегі ннтерфазалың хромосома; 2 —метафазалыңхромосома. Бұл жағдай метафазада ДНҚ-ның 100—500 есеге дейін «тығыздала» (спирализация, коиденсация) түсетінін көрсетеді. Жіпшелердің қабаттасып бүктелу моделі классикалық генетиканың мәліметтерімен де үйлеседі (9, 21-тарауды қараңыз). Алғашқы гипотеза тіпті расталмаған күннің өзінде хромосомадағы қүрылым иерархиясын түсінуге мүмкіндік береді.
Митоз циклінде хромосома жуан-дығы езгеріп отырады. Мысалы, профазада жіқішке хромосомалар жуандай, жинақтала бастайды. Метафазада олар жуандап қысқарған дейін болып керінеді.
Метафаза — пластинкасына тән хромосомалардың қысқарып қалыптасу механизмі қандай? Митозға дайындық кезінде хромосомадағы хромонемалар спиральдану процесіне үұшырайды. Профазаның барысында спиральдану барлық хромосомоға таралады, сөйтіп метафаза кезінде ол ең жоғары сатыға көтеріледі. Сондықтан метафазада хромосомалар өте жинақы болып көрінеді.
Казір спиральданудың екі түрлі болатыны анықталды. Олар ірі және үсақ спиральдану, бүл кездерде жіпшелер орамдары өзара перпендикуляр орналасады. Спиральданудың екі түрі де бір мезгілде жүреді (6-сурет).
Телофазада хромонемалардың деспиральдануы басталады, интерфазада хромонемалар әбден тарқатылған күйде болады. Митоз циклінде хромосомалардың
спиральдану және деспиральдану ерекшелігі заңды процесс (спиральдану циклі, 7-сурет).
Метафазада хромосомаларының кәлемінің үлғаюы тек спиральданумен ғана байланысты емес, конденсациямен — жіпшелердің тығыздалып орналасуымен де, оларды әр түрлі заттардың қоршауымен де, яки кабықша — матрикс түзілуімен де бай-ланысты болады. Матрикс негізінен ядрошық заттарынан тұрады және ол телофазаға дейін сақталады. Онын, ролі әлі толық анықталған жоқ, бірақ бұл кезде хромосомалардың жаңа жас клеткалар арасында рибонуклеопротеидтерді бөлуші ретінде қызмет атқаратыны күдік тудырмайды.
Хромосомалардың ұзына бойына жіктеліп дифференциялануына ерекше назар аудару керек.Профазада хромосомалардың бүкіл ұзына бойына қарайып боялатын дәндер, яки хромомералар байқалатыны жоғарыда ай- 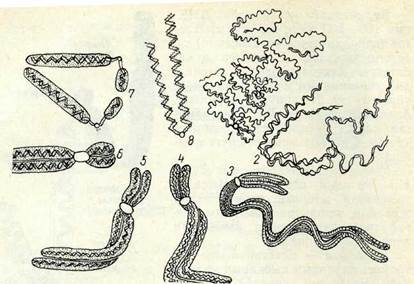
7-сурет. Хромонеманың митоздық циклде спиралдану схемасы:
1 —интерфаза, хромоыемалар әлсіз спиралданған (қалдың спиралдар); 2, 3, 4 —про-фаза, хромоыема спиралдануының күшеюі екі хроматидтің түзілуі; 5 — прометафа-за, төрт жярты хроматидтің көрінісі; 6 — метафаза, ең кушті спиралдануы, мүнда үлкен спиральда, кіші сииральда байңалады; 7 — анафаза; 8 —■ телофаза (жас хро-мосомалардьщ біреуі) хромонеманың деспиралдануы (дөңгелектермен центромералар көроетілген).тылған еді. Олардың әр хромосомада орналасу орны тұрақты және әр хромосомада ол әр түрлі болады. Хромосоманың бұл таспиқ тәріздес жуантық жері ең алдымен спиральданатын хромонемалардын, белігі.
Ол ен, тығыздалған бәлігі болып табылады, сондықтан кәдімгі жарық микроскоптан қарайған туйіршік сияқты болып көрінеді.
Негізгі бояулармен бояп, фиксация жасаған кезде әр түрлі учаскелер түрлі реакция береді. Бір учаскелер жақсы боялады — олар гетерохроматинді, ал екінші учаскелер нашар боялады — олар эухроматинді учаскелер деп аталады.
Гетерохроматинді учаскелер хромосоманың бүкіл ұзына бойына таралған, көбінесе центромераға жақын және хромосомалар ұшында, сол сияқты ядрошықтын, маңында орналасады. Хромосоманың кейбір түрлерінің басым бөлігі гетерохроматиннен тұрады. Гетерохроматинді учаскелер клетканың букіл тіршілік циклі бойы, солардың ішінде интерфазада да, қатты спираль-данған күйде болады, сірә сондықтаы да күштірек боялатын болса керек.
Жіктеп дифференциациялап бояу кезінде байқалғандай, гетерохроматин біртектес емес: одан клетка бөлінуінің бүкіл циклі кезеңдерінің бәрінде хромосомада болатын конститутывті заттар және клетка циклінің бір бөлігінде немесе гомологиялық хромо сомалардың біреуінде ғана кездесетін факультативті заттар табылды. Иитерфазада эухроматинді учаскелер деспиральданады, бұл олардың өте жоғары мотеболистік активтігін көрсетеді. Сірә, интерфаза ядросындағы хроматин тұйірлері, хромссоманың гетерохроматинді учаскелері болуы мүмкін. Қоршаған ортанық әсеріне олар әсіресе тұрақсыз болады, хромосомалардың үзілу жиілігі және олардың қалпына келу қабілеті оларға байланысты болады.
ДНК молекулаларына денатурация — ренатурация белгілі құрылысы бар ДНК жоне РНК молекулаларын будандастыру, басқа әдістер қолдану мынаны көрсетті: гетерохроматиннің эухроматиннен айырмасы оиың спиральдану дәрежесінде ғана емес, химиялық құрамы бойынша да айырмашылығы бар. Гете-рохроматиндегі ДНК. нуклеотидтері қысқа және б£ЛгШ бір ретпен қайталап отырады, көбінесе оларды қайталамалар, репликтер деп атайды, ал ДНК — сателлитті деп аталады. Эухроматин-де ДНҚ нуклеотидтері ұзын, қайталанбайтын бірыңғай ретпен орналасқандығы байқалады (17-тарауды қараңыз).
Хромосоманың үзына бойына дифференциациялануы алып хромосомалардан әсіресе жақсы көрінеді. Дене және жыныс клеткаларыыың кәпшілігінің қалыпты метафазадағы хромосомаларымен салыстырғанда,
алып хромосомалар.100—200 есе ұзын және ондағы хромонемалар да 1000 есе көп болады. Мұндай кү-рылымы бар хромосомалар қос қанатты насекомдар личинкасының дене клеткасындағы ядроға, ішекқарын, малышгий тамырлар, сілекей бездер клеткаларына тән болады, сол сияқты ондай хромосомалар кейбір әсімдіктерден (антиподалар мен синергидтерде) және карапайым организмдерден табылды.
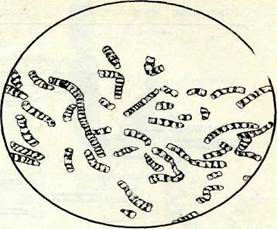
Алып хромосомалардың политендігі, эндомитоз кезінде пайда болады. Бүл жағдайда 2 хромонема жүйелі тоғыз рет екіден еселенгеннен кейін бір-біріне тығыз орналасқан 1000-ға жуық жіпше түзеді. Алып хромосомалардың хромонемалары үнемі біршама және біркелкі деспиральданған күйде болмайды, ол жағдай хромосомалар ұзындығының 100—200 есе ұлғаюына себеп болады. Кәдімгі алып хромосоманы дрозофила личинкаларының сілекей бездерінен (Огозорһііа туысы, 8-сурет) байқауға болады.
Олардың ерекшеліктеріне мыналар жатады. Морфологиясы және келемі бірдей хромосомалар (біреуі әкесінің, екіншісі шешесінің) бірігіп, коньюгациялана (дене конъюгациясы) алады, бұл жағдай алып хромосомалардың жуандығын одан әрі үлғанта түседі.
Екінші ерекшелігі өте көп хромонемалардың хромомералары біріне-бірі жанаса отырып, өзара тығыз орналасады, буылтықтар — дискілер түзеді, олар боялған кезде қарайып боялады және көлденен, сызылған сызықтар сияқты болып көрінеді.
Дискілердің көлемі мен морфологиясы өте өзгеріп отырады, бірақ әр хромосоманың бүл көрсеткіштері түрақты болады және хромосомаларды ұқсатып — ажырату үшін өте жақсы маркер болып табылады. Хромосомалардын, политенділігі дискілер арасынан жақсы көріыеді. Дискілердің құрылысы онтогенезде өзге-
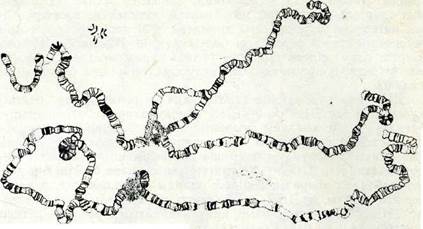
8-сурет. Дрозофиланың сілекей безі клеткалары (алып) жэне ганглия клеткалары
(митоздьтқ) ядросыидағы хромосомалардьщ салыстырмалы көлемі.реті, ол өзгеру хромосоманың қызмет атқаруына байланысты болады, ал ол жөнінде 22-тарауда баяндалады.
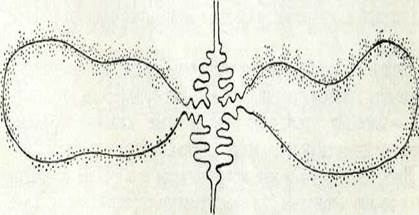
Хромосомалардың нәзік құрылысымен және олардың қызме-тімен танысуға болатын екінші модель «лампа щеткасы» сияқты хромосомалар. Бұл хромосомалардың түрлері әйнек пробирка-ларды жууға арналған ершке үксайды, ертеде онымен керосин лампалардың шыныларын жуатын еді. (I табл., Б). Бұл хромосомалардың жекелеген учаскелері өте созылған және хромосома осіне перпендикуляр симметриялы ілмектер жасайды (9-сурет). Хромосомалардың мұндай күйі балықтың, амфибияның, рептилияның және құстың ооциттерінде кездеседі.
| 9-сурет. «Лампа щеткасы» типтес хромосоманыц жеке ілмегінің құрылыс схе-масы. |
«Лампа щеткаларының» алып хромосомалардан айырмасы— олар политенді болмайды, күшті деспиральданған хромонемалары болады. Өте күшті деспиральдануы ооциттердіқ өсуі кезінде хромосбмалардың метаболистік белсенділігінің артуымен
байланысты деп жобалайды (17, 21-тарауларды қараңыз
 |
| 10-сурет. Жеке нуклеотидтің (жоғарғысы) және ДНК жеке тізбегі- нің құрылысы (төмен-г і с і); ф— фосфор кыпщылының ңалдығы; Д — дезоксирибо-за; Л— пуриндік немесе пиримидиндік негіз. |
Хромосомалардың химиялық құрамы. Хромосомалардың химиялық құрамын зерттеу, олардың негізінде нуклеопротеидтерден (90—92%) тұ-ратынын көрсетті. Нуклеопротеидтер дезоксирибонуклеин қышқылынан (ДНК) және белок гистоннан (немесе протаминнен) тұрады. Хромосомада бұдан өзге РНҚ, біраз мөлшерде кальций, магний, темір т. б. иондарды және кейде РНҚ комплексін түзетін гистонды емес белоктар болады.
ДНК өзінің табиғаты бойынша биологиялық полимер болып табылады, оның құрылымы күрделі құрылысты таспа сияқты. ДНҚ-ның молекулалық массасы 7-Ю6—100• 106Д шамасына жетеді.
ДНҚ молекуласы кезектесіп отыратын мономер бірлігінен дезоксирибо-нуклеотидтерден құрылады. Әр нуклеотидтің құрамына гетероциклді азот-ты негіз (пуринді немесе пиримидинді), дезоксирибоза қанты және фосфор қышқылының қалдығы кіреді (10-су-рет). Басым көпшілік дезоксирибонук-леоидтер құрамына универсалды таралған гетероциклді негіздер: пурин туындылары — аденин мен гуанин және пкримидин туындылары — щітозин мен тимын кіреді.
ДНҚ тізбегіндегі нуклеотидтер арасындағы байланыс қатал біртекті және ондай байланыс фосфор қышқылынын, диэфирі түзілу арқылы, көрші дезоксирибоза қалдықтарының 3 және 5-гидроксилдері арасында пайда болады.
Сонымен, ДНК полимер тізбегі кезектесіп отыратын дезоксирибоза және фосфат қалдықтарынан тузіледі. Осы тізбек қаңқасындағы әр дезоксирибоза қалдыққа бүйір радикалдар — пуриндік немесе пиримидиндік негіздер жалғасқан.
Өсімдіктердің, жануарлардың және микроорганизмдердіқ ДНК-ның химиялық қүрамын зерттеу нәтижесінде әр түрдегі пуриндік және пиримидиндік негіздер өзіндік ерекше таралатындығымен сипатталатындағы анықталды. Әр түрде гуаницитозин мольдер санының аденинтимин мольдері санына қатысы 0,45-тен 2,8
шамасына дейін ауытқып отырады (басқа да алшақ ауытқулар кездеседі). Әр түрдегі ДНҚ макромолекуласында нуклеотидтердің кезектесіп орналасуы әр түрлі, мұның өзінен тұқым қуалаушылыққа тікелей қатысты екені көрінеді.Ал ДНК құрылымын қарастыратын болсақ, бұлардың ғажап
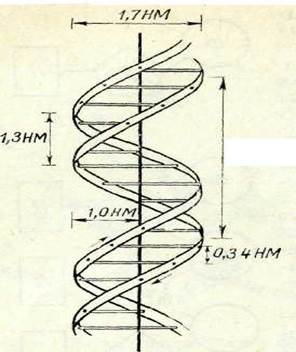
| 13-сурет. ДНҚ кос спира-лыныц кұрылым схемасы (Уотсон — Крик моделі) |
біртектес екенін байқаймыз. ДНҚ-ның бір молекуласында екі полинуклеотидтер тізбегі оң жаққа қарай бұралған екі спираль (бұрандалы баспалдақ сияқты) болып біріккен, бүл кезде екі тізбектің де п-рин және пиримидин негіздері спиральдар бұрандасының аралығындағы кеңістікте болып шығады. Негіздер өзара сутекті байланыстар арқылы жалғасқан. Қалыпты жағдайда бір тізбектегі пурин негіздерге екінші тізбектегі пиримидин негіздегі сәйкес келеді және керісінше атап айтқанда: аденин әрқашанда тиминмен, гуанин цитозинмен байланысты болады. Сәйтіп ДНК-ның екі жіпшесі бірін-бірі өзара толықтырып отырады, ол комплементарлық деп аталады. ДНҚ-ның қос спиральды құрылым схемасы 11-суретте берілген.
Хромосома құрамына РНҚ кіре-ді, ол да ДНК сияқты, полинуклеотид болып табылады. Оның құрамында төрт азоттық негіз болады: аденин мен цитозин, гуанин мен урацил. РНК-да тимин-нің орнын урацил, ал дезоксирибозаны рибоза ауыстырады. ДНК-дан РНК-ның ерекшелігі әдетте оның құрылымы бір тізбекті болып келеді. ДНК-ны клеткадан беліп алудын, жұмсақ әдісін қолданғанда, яки молекуланы бүлдірмей бөліп алу әді-сінде хромосома саны мен альга ДНК молекулалар санының толық сәйкес екені анықталды. Молекулалардың ұзындығы хромосомалардың ұзындығына пропорционал болып шықты. Бұл әрбір хромосомада центромера үзе алмайтын ДНК-ның бір молекуласы болатындығы жөнінде қорытынды жасауға мүмкіндік береді. Мысалы, адамның 1-хромосомасының метафаза кезіндегі ұзындығы 10 мкм, ондағы ДНК молекуласының ұзындығы 16 см
Дегенмен, хромосоманың молекулалық деңгейдегі жалпы конструкциясы әлі айқын емес. Сол сияқты ДНК-ның белокты комплексінщ құрылысы да белгісіз.
Хромосомалар репродукциясы. Тұқым қуалаушылықты түсінуге байланысты генетикадағы негізгі мәселелер мыналар бо лып табылады: 1. Митоздық циклдің қайсы кезінде хромосомалар өздерін ұдайы өндіре алады?
| фазасы; ДНК синтездеу фазасы; 2— постсинтездік фаза. |
| Интерфааа: Я\ — пресинтездік |
2. Өздерін ұдайы әндіріп шығарудың молекулалық механизмі қандай?
Хромосомалар биосинтезінің молекулалық механизмінде, хромосоманың негізгі компоненті ретінде ДНҚ репликациясы маңызды роль атқаратыны анықталды. Дегенмен, ДНК репликациясыныц механизмін білу хромосомалардың екі еселенуі механизмін толық түсіндіре алмайды.
Митоз циклі кезінде ДНК син-тезін зерттеу ол көп клеткалы оо-ганизмдерде интерфазада жүре-тінін көрсетті (12-сурет).
Интерфаза үш кезеңге немесе фазаға бөлінеді. Митоз аяқталысымен басталатын фазабелгіленеді, ол кезде ДНК. синтезделмейді, бірақ клетка құрылымы түзілуіне және келесі бөлінуге қажетті баска өнімдер (олардың ішінде РНК жзйе белок бар) жи-нақталады. Бұл фаза пресинтети-калық деп аталады. Ол ен. ұзақ-қа созылады жоне тұрақсыз болады. Бұл фазаның ұзақтығы бірнеше сағаттан бірнеше тәулікке дейін созылады. Бұдан кейін ДНК-ның синтезделу фазасы (5 фазасы) басталады, бүл кезде клетка ядросындағы ДНҚ мөлшері екі есе ұлғаяды. Бүл фаза қысқа бірнеше минуттан бірнеше сағатқа дейін созылады. 5 фазасы кезінде де РНҚ және белок синтезделеді.
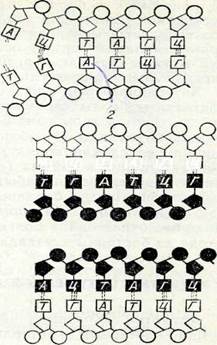
Ең соңында постсинтетикалық фаза (ф2 фаза) басталады, бүл кезде ДНК синтезделмейді, бірақ РНК мен белоктар (әсіресе, ядролық белоктар) синтезі жүреді және келесі митоз үшін энергия жинақталады. Бұл кезде клеткада ДНҚ-ның екі еселенген мөлшері сақталады. Бүл фаза да қысқа, ол епликацияның (қосарланудың) схемасы:бірнеше минуттан бірнеше сағатқа дейін созылады. Осымен интерфаза аяқталады да, митоз басталады, осының нәтижесінде белініп шыққан екі клеткадағы ДНК мөлшері бастапқы клетканікіне тең болады.ДНК-ның екі еселену механизмі қандай? ДНК молекуласының жартылай консервативті екі еселенуі (репликация) деп аталатын схема көпке белгілі (13-сурет). Бұл схема Дж. Уотсон мен Ф. Крик тапқан ДНК-ның құрылым моделімен үйлеседі және генетиканын. мәліметтеріне сәйкес келеді.ДНК-ның жартылай консервативті репликациясы схемасына сәйкес,
ең алдымен аденинтимин, гуанин-цитозин жүптарын түзуші пуриндік және пиримидиндік негіздер арасындағы сутек-тікбайланыстар үзіледі. Осындай үзілістен кейін қос полинуклеотидті тізбек тарқатылады да әр дара тізбек (моноспираль) матрица сияқты, езінің қасында полимеризация жолымен кариоплазмадағы мононуклеотидтерден комплементарлық тізбек түзеді. Осының нәтижесінде бастапкы бірдей екі ДНК молекуласы түзіледі. Сонымен, ДНҚ құрылымының екі жақтылығы, ДНКнуклеотидтершің комплементарлығы, оның репликациялануына қажет-ті аса маңызды шарты болады.ДНҚ-ның репликация процесіне ЛНК-полимераза ферменті қатысанады. Ол ДНК-ның түзіліп жатқан тіз-бегінде З1— ОН — дезоксирибоза тобымен жаңадан келіп қосылатын нуклеотидтің 51—ОН— дезоксирибоза тобы арасында фосфодиэфир байланысының пайда болуын қамтамасыз етеді. Мүндай байланыс түзілу үшін қажетті энергия көзі келіп қосылатын нуклеотидтердің өздері болып табылады. Өйткекі олар энергияға бай қосылыстар — дезоксири-бонуклеозидтрифосфаттар түрінде болады. Нуклеотидтің қосылуы нәтижесінде тағы да анорганикалық пирофосфат бөлініп шығады (14-су-ретті қараңыз). Эксперимент көрсеткендей, ДНК репликациясы жеке фрагменттер (репликондар) түрінде жүреді, олар бүдан кейін біртүтас тізбекке бірігеді (репарацияланады). Бүл процесс те ферменттердің қатысуымен жүреді (бүған лигаза қатысады).Эксперимент жүзінде дәлелденген, ДНК-ның жарты консервативті репликациясы жөніндегі көзқарас, сірә, тұтас хромосомалардың өздерін өндіру процесіне сәйкес келеді. Ол қүбылыс, жоғарыда айтылғандай, интерфазада іске асады. Бүл жағдай Дж. Тейлордың тәжірибелерімен дәлелденеді, ол тәжірибеде авторадиография әдісі қолданылып, ат бұршақ өсімдігінің тамыршасы клеткаларында бастапқы хроматидтер мен жас хроматидтердің тағдыры бірнеше митоз бойы бақыланады. Хроматидке белгі салғаннан кейінгі бірінші митозда түзілген екі хроматидтің екеуі де таңбаланған болып шықты 2). Ал таңбалан-ған тимидині жоқ ортада өткен екінші митозда, әр аналық хромосомадағы екі хроматидтің біреуінде ғана таңба болды, өйткені жаңа репликацияланған хроматидтерге таңба түсуі мүмкін болмай қалды. Бүл зерттеулер мынадай қорытынды шығаруға мүмкіндік берді, хромосома функционалды екі бөліктен түрады, репродукциядан кейін терт бөлік пайда болады және олар ажырап бөлінген кезде жаңа пайда болған жас хромосома құрамында бастапқының бөлігі.
 |
 |
 |
Молекуласының қосарлануының жартылай консервативті механизмін бейнелейтін схема:
1 —ДНК молекуласының бастапңы учаскесі; 2 — азотты негіздердің екі тізбегі ара-сындағы сутекті байланыстардың үзілуі; 3 —ңоршаған орта нуклеотидтерінен (су-ретте — ңара түсті) комплементарлы тізбектердің түзілуі, нуклеотидтер байланысңан кезде пирофосфат бөлініп шығады (ф—ф); 4 —екі жас ДНК молекуласы. Азотты иегіздер әріптермен белгіленген: А — аденин, Т — тимин, Г—■ гуанин, Д— цитозин.
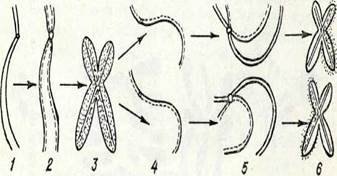
15-сурет: Митоз кезінде бастапқы (тұтасқан сызықпен белгіленген) және репликацияланған (үзік сызықпен белгіленген) хроматид пен хромонема-лар:
1, 2, 3 белгіленген тиминді ортаға енгізгеннен кейінгі бірініші бөлінудің интерфазасьі, профазасы, метафазасы 4, 5, 6 — ортада белгіленген тимин жоқ кездегі екінші бөлінудің сол фазалане жаңа бөлік те болады. Басқа сөзбеи айтқанда, бұл тәжірибелер хромосоманың өзін ұдайы өндіруінің матрицалық, жартьг консервативтік механизмін дәлелдеді. Әр түрлі организмдер хромосомаларын зерттеу осы мәліметтерді дәлелдеді, бұл жағдай жоғары сатыдағы орғанизмдер үшін осы механизмнің уни-версалдылығын көрсетеді.
Авторадиография әдісі, сонымен қатар, мынаны көрсетті: клеткадағы барлық хромосомалар бір мезгілде репликацияланбайды, тіпті әр хромосоманың жеке учаскелері белгілі ретпен репликацияланады (I таблица, В). Хромосоманың түрлі бөліктерінің редупликациясындағы мұндай асинхрондылық — хромосоманың ұзына бойына (дифференциацияланып) жіктелгендігінін. бір дәлелі болып табылады және ондай жіктелу, олардық қызметімен тікелей байланысты болады. Барлық айтылғандардан мынадай маңызды қорытынды шығады: көбею процесі кезінде бөлінуден пайда болған хромосомалар саны жаңа клеткаларда да бастапқы клеткадағыдай болады.
3. КАРИОТИПТІҢ ТҮР ЕРЕКШЕЛІГІ
Кариотип. Жоғарыда айтылғандай, хромосоманы зерттеу кезінде клетканың экваторлық пластинкасында оларды сынауға болады, ал хромосомалардың морфологиясын, құрылымын және шамасын талдау олардың өзара ұқсастығы мен айырмасын дәл анықтауға мүмкіндік береді.ІБір түрге жататын организмнің әр түрлі дене тканьдері клеткаларының хромосомаларын зерттеу мынаны көрсетті: әр түрдің хромосома саны және оның құраліы оның езіне ғана тән болады. Жануарлардың немесе өсімдіктердін, белгілі бір систематикалық тобының дене клеткасындағы хромосомалар жиынтығын кариотип деп атайды.
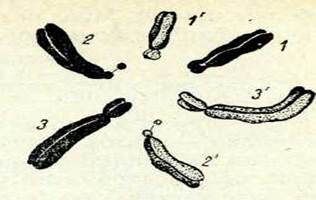 |
| 17-сурет. Сгеріз сарііІагіз клеткасында метафаметафаздық хромосомалардың диплоидты жиынтығы (2н = 6). Гомологиялық хромосомалар бірдей цифрлармен белгіленген. |
. 
Егер кез келген дене клеткасыбың кариотипіне зейін салып қараса, әр хромосома жұп, морфологиясы, құрылымы және шамасы бір-бірінен айнымайтынын байқауға болады.
Хромосоманың мұндай жуптығы мынадан шығады: ұрықтану процесіндегі зигота түзілу кезінде оған хромосомалардың жартысы сперматозоидтан. ал жартысы жұмыртқа
клеткадан келіп қосылады. Өсіпжетілген жұмыртқа клеткаларындағы хромосомалар санын гаплоидты деп атайды және оны п әрпімен бел-гілейді. Көп клеткалы организмніңдене клеткасындағы хромосомалар саны жыныс клеткасыныкінен екі есе көп болады, оны диплоидты деп атайды және 2 п әрпімен белгілейді. Морфологиясьг, құрылымы жоне үлкендігі бірдей, бірақ шыққан тегі әр түрлі (біреуі шешесінікі, екіншісі әкесінікі) жүп хромосомаларды гомологиялық деп атайды.,
17-суретте кәдінің (Сгеріз соріііагіз) хромосомасының диплоидты жиынтығы берілген, онда гомологиялық жұптар көрсетілген.
Кейбір жануарлар мен өсімдіктер хромосомаларының диплоидты санының мысалдары «Генетика жөнінен практикалық сабақтарына басшылықтың» II қосымшасында берілген1.
Кариотиптегі хромосомалар саны жануарлар мен өсімдіктер-дің қүрылыс деңгейіне байланысты болмайды: жоғары дәрежедегі организмдерге қарағанда ең қарапайым организмдерде хромосома саидары көп болуы мүмкін, не керісінше де болады. Же«ке жағдайларда хромосома саны мен морфологиясы жеке организм түрлерінің филогенетикалық туыстығының белгісі де бола алады. Кариосистематика осы принципте жасалады.
Біз әр организм түрінің клеткасында хромосома саны мен формасының тұрақтылық заңы бар екендігін айтқанмен, мүндай тұрақтылық салыстырмалы екендігін ескеру керек.
Тіпті бір организмнің әр түрлі тканьдеріндегі хромосомалар саны, атқаратын функциясына байланысты әр түрлі болуы мүмкін. Мысалы, жануарлар бауырының клеткаларында хромосомалар саны екі жиынтықтан көп (яғни 4п, 8п) болып та кездеседі.
Кейбір өсімдік түрлерінде хромосомалардың саны гаплоидты санға еселенген к
өп болып келуі де мүмкін. Мысалы, қара бидайда (Зесаіе сегеаіе) хромосома саны 2п—14 немесе 28 (4п) 1 Здесь и далее имеется в виду «Руководство к практическим занятиям по генетике» К. В. Ватти и М. М. Тихомировой (М., Просвещение, 1979),
олуы мүмкін. Мұндай кезде екі форманың да қара бидай түрі-не тән барлық белгілері сақталады.Сонымен, митоз нәтижесінде бір клеткадан екі жас клетка түзіледі.
Жоғарыда айтылғандай, клеткакың негізгі бөлігі — хромосомалар — митоз кезінде репродукцияланады, бүл кезде пайда болған екі сіңлілі хроматидтер бірінен-бірі айнымайтын үқсас болады. Ал ахроматин ұршығы осы екі хроматидті теңдей етіп екі жас клеткаға бөліп жібереді, сөйтіп, пайда болған жас клеткадағы хромосома саны бастапқы клетканың хромосомалары санындай болады.
Казіргі кезге дейін ең жаксы зерттелген органоидтар — плас-тидтер мен митохондриялар. Олар ДНК, РНК, ферменттерден тұрады. Олардың ДНК-сы қос тізбекті, сақиналанып түйіскен, молекулалық массасы үлкен болады. Органоидтар ДНК-сының нуклеотидтік құрамы ядроныкінен езгеше. Репликация ядро ДНК-сымен әр түрлі мезгілде жартылай консервативтік әдіспен іске асады. Жоғарыда айтылғандай, пластидтер мен митохон-дриялардың қүрылысы ете күрделі болады, олардың биогенез процесі, яғни қалыптасуы әлі жеткілікті зерттелмеген. Репродукция процесі де нашар зерттелген.
Органоидтардың жаңадан түзілетін клеткалар арасында бөлінуі кездейсоқ, бірақ олар азды-көпті бірдей бөлінеді. Органоидтардың теңдей болып бөліну механизмінің жоқтығы, сірә, олардың санының кептігіне және бірінің орнын екіншісі баса алатындығына байланысты болса керек.
Сонымен, жыныссыз көбеюдің цитологиялық негізі митоз болып табылады, соның нәтижесінде бір клеткадан үқсас екі клетка түзіледі.
3-тарау. ЖЫНЫСТЫ КӨБЕЮДІҢ ЦИТОЛОГИЯЛЫҚ НЕГІЗДЕРІ
Жынысты көбею деп ұрықтанған жұмыртқа клеткасынан — зиготадан, яғни аналық пен аталық жыныс клеткаларының қосылуынан ұрпақтың пайда болуы мен дамуын атайды.
Жануарлар мен өсімдіктерде жыныс клеткаларының жетілу жолы, сол сияқты ұрықтану процесі әр түрлі болады, бірак барлық жағдайда олардын, негізіне ұқсас механизмдер жатады. Өсімдіктер мен жануарлар жыныс клеткалары дамуына ең тән
процесс мейоз болып табылады.
МЕЙОЗ
Мейоздың фазалары. Жыныс клеткалары даму процесінде бір ізді екі бөлінуден түратын мейозға ұшырайды: оның бірін-шісі кәдімгі хромосома санын екі есе азайтатын редукциялық (диплоидты клеткалар гаплоидты 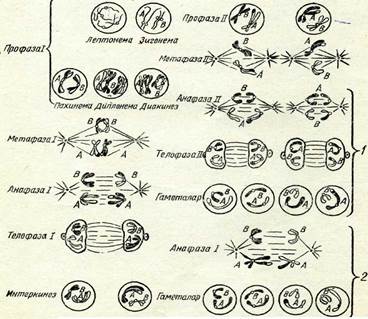
18-сурет. Мейоз схемасы.
| эквациалық (теңестіруші) бөліну, бұл кезде клеткалар хромосомаларының гаплоидты жиынтығын сақтайды.Мейоз циклі бір ізді бірқатар фазадан тұрады, ол кезде хромосомалар заңды өзгерістерге ұшырайды. Бірінші белінуте қа-тысты фазаларды I рим цифрымен белгілеу, екінші бәлінудің фазаларын II рим цифрымен төмендегідей белгілеу қабылдан-ған: |
—
Интерфаз
Профаза I лептонема зигонема пахинема диплонема диакинез Метафаза I Анафаза I Телофаза I
Интеркинез
Профаза II
Метафаза II
Анафаза II
Телофаза II
А шәне В — гомологияльщ хромосомалардың түрлі жұптары: 1 және 2 — гаме-талар тузілу кезінде гомологиялың емес хромосомалардың комбинациялануы-ның мүмкін болатын екі варианты.
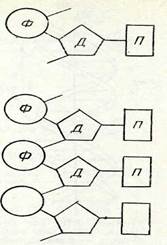
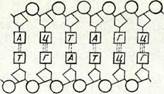
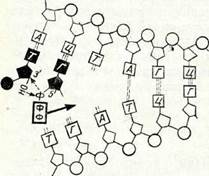
18-суретте және II таблицада (Л) мейоз схемасы берілген. Шыққан тегі әр түрлі (әкесінен және шешесінен) гомологиялық хромосомалар мұнда ақ және қара түспен белгіленген. Мысал ретінде гомологиялық емес екі жұп хромосомалары бар (2п = 4) клеткалар алынған. Мысалы, гаплопаппус (Наріорарриз ^гасі-Ііз) дегеы өсімдіктің, астық тұқымдас өсімдіктің (2іп£егіа Ьіе-Ъегзіеіпіапа) және қан сорғыш шыбындардың бір түрінің (Еизіггшііит зесигііогте) хромосомалар саны осындай болады. 11 профаза кезектесіп келетіи бірнеше стадиядан тұрады. Лептонема стадиясы кезінде интерфазалық ядроның торлы қүрылымы жойылады да, жіңіщке жеке жіпшелер көріне бастайды, олар хромосомалар болып табылады. Жарықтық микроскоптан көрінетін жіпшелердің саны диплоидты болады. Лептонема стадиясы кезінде хромосома жіпшелері ең кемі екі еселеніп жұптанған болатыны электрондық микроскоп көмегімен анықталды, бүл жағдай хромосомалардың екі еселенуі интерфаза кезінде болып кеткенін көрсетеді. Бұл фаза кезінде хромосомалардың спиральдануы әлі жеткіліксіз болғандықтан және олардың жартылары біріне-бірі тығыз жанаса орналасқандықтан, хромосомалардың жұптанған табиғаты жарықтық микроскоптан көрін-■ бейді.
Келесі стадия — зигонемада гомологиялық хромосомалар ұқсас учаскелері арқылы біріне-бірі тартыла бастайды. Олардың жүптанып қосылуы кәбінесе екі шетінен (кейде центромера-лардан) басталады. Жақындасу бір нүктеден басталады да,хромосоманың бүкіл үзындығына таралады. Гомологиялық хромосомалардың мүндай өзара тартылысын коньюгация немесе синапсис деп атайды.
Молекулалық биологияның мәліметтеріне сүйенген қазіргІкөзқарастар бойынша, гомологиялық хромосомалар коньюгациясының негізінде алғашқы қүрылымдары бірдей ДНК молекулаларының бірін-бірі «тануына» байланысты болады. Зигонемада шағын ғана мөлшерде (0,3%) ДНК синтезделетіні авторадиография тәсілімен анықталады; мүндай синтездің нашарлауы хромосомалар коныогациясының бүзылуына әкеліп соғады, сондықтан синтезделетін
ДНҚ коньюгацияның жүруінде белгілі қызмет атқарады деп болжауға мүмкіндік береді.г—Хромосомалар коньюгацияланғаннан кейін, олардың түйісіп| байланысуы синаптонемальды комплекс деп аталатын субмик-роскоптьіҚчДеңгейде үсталып түрылады, бүл өте күрделі, хромосомаларды езара тартып, оларды бірінен-бірі 100 ым қашықтықта үстап түрады.
Хромосомалар коньюгациялары аяқталған, ал олардың спиральдануы ары қарай созылуы нәтижесінде хромосомалардың қысқаруы және жуандауы жалғасатын стадия пахинема деп аталады. Гомологиялық хромосомаларда спиральдану процесі бір мезгілде жүреді. Бүл кезде гомологиялық хромосомалардың әрқайсысының жүптанған қүрылысы жақсы көрінеді. Мүнда әр хромосома екі сіңлілі хроматидтен түрады, оларды бір центромера біріктіріп ұстап тұрады. Коньюгацияланған гомологияльіл, екі хромосома төрт хроматидтен құралған бивалент түзеді. Го-мологиялық екі хромосоманың хроматидтерін сіңлілі емес (не-сестринские) деп атайды. Төрт хроматид тетрада фигурасын түзеді. Хромосоманың нәзік құрылысын зерттеуге осы стадия қо-лайлы болады.
Пахинема стадиясының сонында және келесі стадия — диплонемада (18-суретті қараңыз) хромосомалар ширатылып бірі-не-бірі оралады. Гомологиялық хромосомалардың сәйкес учаскелері бірін-бірі итере бастайды. Бірін-бірі итеру, бірдей зарядталған денелер сияқты, центромерадан басталады да, екі шетіне таралады. Сөйтіп зиготена стадиясында болған қубылысқа кері процесс басталады. Хромосомалардың ажырауы кезінде олардың ширатылуы тарқайды. Осы көрсетілген себептерге байланысты «х» тәріздес фигура түзіледі, ол хиазма деп аталады.Хиазмалардың саны хромосомалар шетіне ауысатындықтан біртіндеп азая береді (хиазмалардың терминализациясы). Егер клеткадағы гомологиялық хромосомалардың жұбындағы екі хромосомалардың морфологиясы екі турлі болса, гомологиялық хромосомалар өзара учаскелерін алмасады деп аталатын құбылыс байқалады. Бұл жүгеріде көрсетілген (9-тарауды қараңыз).Авторадиография әдісі пахинема стадиясында шағын мөлшерде (0,1%) ДНК синтезделетінін көрсетті. Гомологиялық хромосомалар учаскелерінің өзара алмасу процесіне осы ДНК-ның қатысы бар деген болжау бар.Профазаның диакинез деп аталатын соңғы стадиясында хромосомалар ең күшті спиральдануға байланысты қатты қысқарып, жуандайды. Биваленттер оқшауланады, клеткада оларды санауға болады. Олардың
саны гаплоидқа тең болады. Бүл ста-дияда ядрошықтар мен ядро қабығы жойылады. Осымен I профаза аяқталады. Бүдан кейін гомологиялық хромосомалар центромералары бөліну ұршығының экваторы кеңістігіне бағытталып ориаласа бастайды. Бүл I метафазаға сәйкес келеді.
I анафазада биваленттердің гомологиялық хромосомалары ажырап қарсы полюстерге

