 2015-06-28
2015-06-28 4575
4575Жануарлар гаметогенезінің ерекшеліктері. Жануарлардың жыныс клеткасы дене клеткалары сияқты эмбрионалдық клеткалардан пайда болады («Генетиканың практикалық сабақтарына басшылықтағы» 5-суретті қараңыз).
Онтогенезде оқшауланатын, ал кейіннен одан жыныс бездері мен жыныс клеткалары дамитын бастама клеткалар ұрық жолы деп аталады. Ұрық жолының оқшаулануы әр жануарда онтоге-нездің әр кезеңінде, бірақ бәрінде де ол біршама ертерек өтеді. Өзіндей ұрпақ ендіруге және тұқым қуалаушы информацияларды беретін жыныс клеткалары ерте қалыптасып маманданады деп айтуға болады.
Бастама клеткалар бірнеше қайтара бөліну арқылы гониальды клеткаларды — гонийлерді түзеді. Алғашқыда екі жыныс особь.тарында да гонийлер ұқсас болады, одан ары олар жікте-ліп, еркек организмде сперматогонийге, ұрғашы организмде оогонийге айналады.
Олар бірнеше рет митоздық жолмен бөлінеді, бүл кезде хромосомалар жиынтығының диплоидты қалпы сақталады, бірақ шамалы кішірейіп бөлінуі тоқталады. Енді клеткалар өсу кезе-ңіне өтіп, келемі ұлғая бастайды. Хромосомалар жиынтығы диплоидты дамудың бүл стадиясында әлі пісіп жетіле қоймаған аталық. жыныс клеткаларын бірінші қатардағы сперматоциттер (I сперматоцит) деп, ал аналық жыныс клеткаларын бірінші қа-тардағы ооциттер (I ооцит) деп атайды. Аналық және аталық жыныс клеткаларының пісіп жетілу процесінде айырмашылық болады.
Сперматогенез. I сперматоцитте мейоз басталады. Жануар-ларда мейоздық бөлінуді пісіп жетілу бөлінуі деп те атайды. Өсу кезеңінің езінде-ақ I сперматоцитте мейоздың профазасына тән езғерістер басталады. Пісіп жетілу кезеңіндегі бірінші бөлінудің (редукциялық) нәтижесінде екінші қатардағы сперматоциттер (II сперматоцит) түзіледі. Олар гаплоидты болады. Пісіп жетілу кезіндегі екінші белінуден (эквациялық) кейін әр II спер-матоциттен екіден клетка түзіледі. Ол клеткалар сперматидтер деп аталады. Сонымен, диплоидты бір клетка — I сперматоциттен мейоздық жолмен екі рет бөліну нәтижесінде гаплоидты төрт сперматид түзіледі.
Жетіліп, қалыптасу кезінде сперматидтердің сперматозоидтарға айналу процесін
спермиогенез деп атайды. Ядро мен цитоплазманың барлық элементтері осы процеске қатысады. Пісіп жетілген сперматозоидтың басы, орта бөлігі (мойны) және құйрығы болады.
Сперматид ядросы сперматозоидтың басына айналады. Цитоплазмалық органоидтар структуралық жағынан өзгеріске ұшырап, әр түрлі органдарға айналады, ол органдар сперматозоидтық қозғалысын және оның жұмыртқа клеткасына енуін қамтамасыз етеді. Сперматозоидтағы цитоплазма өте жұқа қабат түрінде болады.
Сперматозоид ядросының химиялық құрамы басқа ткань клеткаларының ядросына ұқсас, бірақ кейде онда гистонның орнына протамин болады. Сперматозоид ядросындағы хромосома саны гаплоидты болғандықтан, мұндағы ДНК мөлшері де диплоидты клеткалар ядросыныкінен екі есе аз болады.
Электронды микроскоп көмегімен сперматозоид ядросының құрылысының кристалды болатыны анықталды, мұнда Дезокси-рибонуклеопротеидтің молекулалары параллельді орналасуын қамтамасыз етеді. Сперматозоидтың мұндай күйі тұқым қуалаушы информацияны сақтап жеткізу үшін бейімделгендік болуы мүмкін, өйткені сперматозоид еркек организмінен басқа жерде әр түрлі сыртқы әсерлерге ұшырайды. Шындығында зерттеу пісіп жетілген және пісіп жетілмеген жыныс клеткаларының сыртқы орта әсеріне сезімталдығы әр түрлі болатынын көрсетті (13-тарауды қараңыз).
Морфологиясы жағынан сперматозоидтар алуан түрлі және әр түрдің езіне тән болады.
Оогенез. Аиалық жыиыс клеткаларының — жүмыртқа клеткалары — дамуын оогенез деп атайды. Бұл негізінен сперматогенезге ұқсас, алайда айтарлықтай айырмасы да бар.
Біріншіден, бірінші қатардағы ооциттердің (I ооцит) өсу стадиясы I сперматоцидтердікіне қарағанда ұзақ болады, өйткені өсу кезінде оцитте — болашақ жұмыртқа клеткасында — қоректік заттар жиналады. Осы стадия кезінде кейбір жануарларда «лампа щеткалары» сияқты хромосомалар пайда болады.
Екіншіден, мейоздық жолмен екі рет белінгеннен кейін әр-бір I ооциттен төрт оотид түзілгенімен, соның біреуі ғана (одан әрі жұмыртқа клеткасы) есіп-жетіліп, үрықтана алады. Гаплоидты хромосомалар жиынтығы бар қалған үш оотидте цитоплазма қоры жеткіліксіз болады, олар тіпті өз алдына жеке клетка болып оқшаулана алмайды. Олардың қалыптасып түзілуі былай жүреді. Пісіп жетілудің бірінші бөлінуінен кейін, II
ооциттен баска, бірінші полярлық дене бөлініп шығады; оны басқаша бағыттаушы немесе редущиялық дене деп атайды. Полярлық дене бөлініп, екі оотид түзеді. Пісіп жетілудің екзнші бәлінуі нәтижесінде жұмыртқа клеткасы және екінші полярлық дене, яғни үшінші отид түзіледі.
Сонымен, оогенез процесі кезінде екірет мейоздық жолмен
бөліну нәтижесінде пайда болатын төрт клетканың біреуі ғана жұмыртқа клеткаға айналады.
Бұның жыныстық жолмен көбеюдегі заңдылықтарды түсінуде үлкен маңызы бар. Жоғарыда айтылғандай, мейоз процесі әкелері мен шешелерінің хромосомаларының түрлі комбинациялануымен клеткалар түзіледі, ал оогенез нәтижесінде тек бір клетка ғана, яки пайда болған барлық комбинациялардық біреуі ғана тіршілік ете алады.
3. ӨСІМДІКТЕР СПОРОГЕНЕЗІ МЕН ГАМЕТОГЕНЕЗІ
Өсімдіктер гаметогенезінің ерекшеліктері. Өсімдіктерде жыныс клеткаларының қалыптасу процесі екі кезеңге бөлінеді: 1-кезең — спорогенез — гаплоидты клеткалар — споралар түзілуімен аяқталады; екінші кезеңге — гаметогенез — жетілген гаметалардың түзілуі кіреді.
Өсімдіктерде микроспоралар немесе тозаң түйірлері түзілу процесі микроспорогенез деп, ал мегаспоралар (немесе макроспоралар) түзілу процесі мега немесе макроспорогенез деп аталады.
Өсімдіктердегі гаметогенез процесі негізінде жануарлардағы сондай процеске ұқсас болады, бірақ біршама басқа жол-мен өтеді. Өсімдіктерде ұрықтық жол, яғни жыныс клеткаларының окшаулануы ертерек болмайды («Гейетика жөнінен практикалық сабаққа басшылықтағы» 6-суретті қараңыз).
Микроспорогенез және микрогаметогенез. Жабық тұқымды өсімдіктерді жалпы мысал етіп алып, микроспорогенез бен микрогаметогенезді қарастырайық. Жас тозақдықтың субэпидермальды тканінде археспория деп аталатын клеткалар пайда болады, олардың әрқайсысы бірнеше рет бәлінуі нәтижесінде тозаңнық аналық клеткасына айналады да мейоздың барлық фазасынан етеді.
Екі рет мейоздық бөліну нәтижесінде гаплоидты терт микроспора түзіледі. Олар төрт-төрттен жатады, сондықтан -ралар тетрадасы деп аталады. Пісіп жетілу кезінде тетрадалар жеке микроспораларға бөлінеді.
Осымен микроспорогенез аяқталады.
Бір ядролы микроспора түзілісімен микрогаметогенез басталады. Микроспоранын, митоздық бірінші бөлінуі нәтижесінде вегетативті және генеративті клеткалар түзіледі. Бұдан ары вегетативті клетка мен оның ядросы бөлінбейді. Онда қоректік заттар жиналады, ол заттар кейіннен генеративті клеткалардың бөлінуін және аналық жыныс мүшесінің бағанасындағы тозан түтігінің өсуін қамтамасыз етеді.
Құрамында цитоплазмасы аз болатын генеративті клетка тағы бәлінеді. Мүндай бөліну тозаң түйірінде немесе тозаң түтігінде орындалуы мүмкін. Осының нәтижесінде аталық екі жыныс клеткасы түзіледі, олардын, жануарлар сперматозоидынан айырмасы қозғала алмайды және спермия деп аталады.
Сонымен, гаплоидты хромосома жиынтығы бар бір спорадан митоз жолымен екі рет бөліну нәтижесінде үш клетка түзіледі. Олардың екеуі — спермия, біреуі — вегетативті клетка. /^ Мегаспорогенез бен мегагаметогенез. Жас түқым бүршігінің субэпидермалық қабатында археспория клеткасы дараланып рқшауланады, көбінесе ол біреу ғана болады. Археспория клеткасы өсіп, мегаспораның аналық клеткасына айналады.
Мегаспора аналық клеткасының мейоздық жолмен екі рет бөлінуі нәтижесінде мегаспоралар тетрадасы түзіледі. Тетраданың әр клеткасы гаплоидты болады. Ал осы тетраданың бір клеткасы ғана ары қарай дами алады, қалған үшеуі дегенерацияланады (дамып жетілудің моноспоралық типі). Бүл клеткалардың тағдыры, жануарлардың жұмыртқа клеткасы пісіп жетілуі кезіндегі редукциялық денелердің тағдырына ұқсайды.
Келесі кезеңде мегагаметогенез іске асады. Ларамды күйінде қалған мегаспора ары қарай өсе береді, бұдан оның ядросы митоз жолымен бірнеше рет бәлінеді. Сонымен клетканың өзі белінбейді, ол ұрық қалтасына айналады.
Әр түрлі өсімдіктерде митоздар саны бірден үшке дейін өзгере алады. Қөпшілік өсімдіктерде (жабық тұқымдыларының 70%) бөліну үш реттен болады, осының нәтижесінде бірдей сегіз ядро түзіледі. Осындай бөліну уақытында ядролар полярлық күйде болады, олардың тертеуі микропильге (спермиялардың енетін жері) жақын жатады, ал қалған төртеуі халаза деп аталатын қарама-қарсы белігінде орналасады. Ары қарай ядролар дербес клеткаларға айналып оқшауланады, ол клеткаларда едәуір мөлшерде цитоплазма болады.
Микропиледе орналасқан төрт клетканын, үшеуі — жұмыртқа клеткасымен синергидтер деп аталатын екі клетка — жұмыртқа аппаратын қүрады.Синергидтер ұрықтану кезінде көмекші қызмет атқарады, көп ұзамай олар ыдырап жойылады. Төртінші ядро ұрық қалтасының ортасына кетеді, сол арада халазалық шеттен келген бір ядромен қосылады. Орталық белікте құйылысқан екі гаплоид ядродан бір диплоидты ядро, екінші кезектегі, яки үрық қалтасының орталык ядросы түзі-леді. Ұрық қалтасынын, халаза шетінде қалған үш ядро клеткаға айналып оқшауланады; олар антиподтар деп аталады. Антиподтар, синергидтер сияқты, зиготаның дамып жетілуі кезінде кемекшілік қызмет атқарады да, кеп ұзамай, ыдырап бұзы-Сонымен, митоздық жолмен үш рет бөліну вәтижесінде ұрық қалтасында гаплоидты және бірдей 8 ядро түзіледі, солардың біреуі ғана жүмыртқа клеткасына айналады.
- Біз қарастырып отырған бір мегаспорадан сегіз ядролы ұрық қалтасының түзілу схемасы көп таралған. Бірақ әсімдік-тердің әр түрлі топтарында бүл процесс алуан түрлі болып өтеді.
Өсімдіктер мен жануарлардың филогенездік ажырауы (дивергенция) клеткалы құрылым түзілуінің өте ерте кезінде болды. Соған қарамастан жануарларда және өсімдіктерде жыныс клеткаларының шсіп жетілуш салыстыру, олардың арасындағы толық параллелизмді көрсетеді.
4. ҰРЫҚТАНУ
Үрықтанудың мәні. Аталық және аналық жыныс клеткаларының ядролары қосылу (кариогамия) арқылы жұмыртқаның дамып жетілуіне жағдай тууын ұрықтану деп атайды. Ұрықтану — кері жүрмейтін бір беткей процесс: Бір рет ұрықтанған жұмыртқа клеткасы қайтадан ұрықтанбайды. Сингамия (аталық және аналық жыныс клеткаларының қосылып бірігуі) мен кариогамия ұрықтанудың негізгі мәнін білдіреді.
Жануарлардағы ұрықтану. Жануарларда болатын ұрықтану процесін бірнеше фазаға бөлуге болады. Жұмыртқа клекасының кез келген бір нүктесіне сперматозоидтың бекінуі немесе микропиле арқылы оған енуі — бірінші фазаның басталуы. Сперматозоид басының жұмыртқа клеткамен түйісуі химиялық реакциялар тізбегінің бастамасы болып табылады. Бүл фазаны жұмыртқаның активтену фазасы деп атайды.
Жұмыртқа клеткасына бір сперматозоид (моноспермия), ал кейбір жануарларда бірнеше сперматозоидтар (полиспермия) енгеннен кейін үрықтану
процесінің екінші фазасы-басталады. Жүмыртқа клеткасының ішіне енген сперматозоид аналық ядромен қосылып бірігуге «дайындалады», ол митоз бойынша жүреді: сперматозоид ядросы біртіндеп ісінеді де интерфаза кезіндегі ядроға ұқсайды. Мұндай ядро аталық про~ нуклеус деп аталады. Сперматозоид ядросымен қосылып бірігуге дайын болған (мейоздың барлық фазасынан өткен) жұмыртқа клеткасының ядросын аналық пронуклеус деп атайды.
Сперматозоидтың жүмыртқамен түйісу және оның ядросына енуі кезінде әр түрлі жануарларда жұмыртқа бөліне отырып жетілудің әр түрлі бөліну стадиясында болуы мүмкін. Мейоз аяқталғаннан кейін ғана, яғни аналық пронуклеус түзілгеннен кейін ғана кариогамия іске асады.
Үрықтану актісінде гаплоидты екі пронуклеус бір ядро болып бірігеді де, зигота түзеді. Осы мезгіл жыиысты көбею процесінін кульминациялық шегі болып табылады. Алдыңғы үр-пақтағы мейозда ажырап кеткен гомологиялық хромосомалар кариогамия нәтижесінде зиготаның бір ядросына қайтадан бірігеді. Жыныстық көбеюде хромосомалардың диплоидты жиынтығы осылай калпына келеді.
Өсімдіктердегі ұрықтану. Өсімдіктердегі үрықтаиу процесі негізінен жануарлардағыларға ұқсас, дегенмен бірқатар ерекшеліктері де бар.
Бұрын айтылғандай, ёкі спермия түзілумен микрогаметогенез аяқталады, ол екі спермия нетозаң дәнінде, немесе тозаң түтігінде тозаң дәиінің өсуі кезінде қалыптасады. Тозаң түтігі өсіп, ұрық қалтасыыың микропилесіне дейін жетеді де жұмыртқа аппаратымен — жұмыртқа клеткасымен және синергидтер-
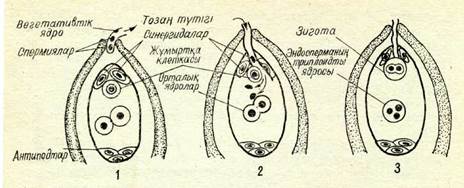
19-сурет. Өсімдіктегі қос қабат ұрықтану схемасы:
1 — тозяң түтігінің ұрың ңапшығына енуі; 2 —■ тозаң түтігіндегі заттың ұрықтапшыныға құйылуы; 3 — ұрықтанғаннан кейінгі ұрынапшығы мен түйіседі (19-сурет, 1). Тозаң түтігінің ұшы синергидтермен түйіскеннен кейін
тозаң түтігі жарылады да синергидтер бүлініп ыдырап кетеді. Тозаң түтігімен жүріп, жылжып келе жатқан генеративті екі ядро — спермиялар — түтік өсе түсіп, жарылғаннан кейін, оның барлық заттарымен бірге ұрық қалтасының ішіне құйылады (19-сурет, 2). Ұрық қалтасына енген екі спермияның біреуі жұмыртқа клеткасының гаплоидты ядросымен қосылып бірігеді (19-сурет, 3). Спермия ядросының жұмыртқа клеткасы ядросымен қосылып бірігуі өсімдіктердегі нағыз ұрықтану болып табылады. Ұрықтанған жұмыртқа клеткасында — зиготада — хромосомалардың диплоидты жиынтығы қалпыыа келеді. Зиготадан түқым ұрығы өсіп-жетіледі.
Өсімдіктерде де, жануарларда да спермиялар ұрық қалтасына кірер алдында аналық ядро әр түрлі стадияда болуы мүмкін, ал кариогамия мейоз аяқталғаннан кейін ғана орындалатынын атап өту қажет.
Жабық түқымды өсімдіктер тұқымында ұрықтан өзге, эмбриональдық қосымша орган — эндосперм өсіпжетіледі, ол үрықтың азық қоры болып табылады. Эндоспермнің дами бастауы екінші рет ұрықтанудан басталады. Тозаң түтігіндегі екінші спермия ұрық қалтасына түскен соң, ұрық қалтасындағы орталық клетканың диплоидты ядросымен қосылып бірігеді. Бүл кезде хромосомалар саны үш есе көп клетка түзіледі: оның құрамында аналық организм хромосомаларының екі есе жиынтығы және аталық организм хромосомаларының бір есе жиынтығы болады (19-сурет, 3). Бір спермияның жүмыртқа клетка-сымен, екінші спермияның орталық клетка ядросының құйылып қосылуын қосарынан ұрықтану деп атайды. Қосарынан ұрықтануды 1898 жылы біздің отандасымыз С. Г. Навашин ашты.
Жануарлар мен өсімдіктердегі ұрықтану процесінін, жалпысипаты осындай болады.
Моноспермия, полиспермия, таңдаушылық жэне селективті
ұрықтану. Жануарлардың және өсімдіктердің бір жұмыртқа клеткасына келетін сперматозоидтар мен тозаң түйірлерінщ саны орасан көп болғанымен, ұрықтану процесіне әдетте бір ғана сперматозоид пен бір тозаң түйірі қатысады. Ұрықтанудың мұндай түрі моноспермиялы ұрықтану деп аталады. Мұндай үрықтану жануарлардың және өсімдіктердің көпшілігіне тән болады.
Моноспермиялы үрықтануды бірқатар механизмдер бақылайды.
Дегенмен, жануарларда жұмыртқа клеткасының цитоплаз-масына бірнеше сперматозоид кіреді. Жұмыртқа клеткасының цитоплазмасына бірнеше сперматозоидтың ену құбылысыь полиспермия деп атайды. Полиспермия ұлулар, тікен терілілер, насекомдар сияқты омыртқасыздарда кең таралған. Ол құбылыс балықтар (акула тәрізділерде заңды түрде), амфибиялар, рептилиялар және құстар сияқты омыртқалыларда да кезде-седі.
Сүт қоректілерде полиспермия әдетте сирек кездеседі (1—2%); бір тесіктілер бүл ережеге бағынбайды, оларда әдетте осы құбылыс кездеседі. Полиспермия құбылысы өсімдіктерде егер ұрық қалтасына бірнеше тозаң түтігі еніп кеткенде байқалады. Қызылшада, мақтада, қарақұмықта, темекіде және басқа өсімдіктерде полиспермия байқалды.
Алайда полиспермия құбылысында жүмыртқа клеткасына бірнеше сперматозоидтың еніп кеткеніне қарамай, аналық нуклеуспен бір ғана аталық нуклеус қосылады. Басқалары жойылып кетеді. Полиспермді кариогамия, яғни бірнеше нуклеустер-дің құйылып қосылуы әдетте, кездеспейді. Өсімдіктерде қосымша спермиялар жүмыртқа клеткасы ядросымен емес, ұрық қалтасындағы басқа ядролармен (синергидтермен немесе антиподтармен) қосылып бірігу құбылысы байқалғаны белгілі. Мұн-дай жағдайда бір ұрық қалтасынан бірнеше ұрық (полиэмбриочия) пайда болады.
Жұмыртқа клеткасы цитоплазмасына бірнеше спермияның енуі және оның біреуінің ғана жүмыртқа клеткасының ядросымен қосылып бірігуі, бұл процесс кездейсоқ механикалық процесс емес деген ойға көптен бері итермеледі. Кариогамия процесінде «таңдаушылық», яғни аналық пронуклеустің белгілі бір аталық пронуклеуісті «таңдап», онымен қосылуы мүмкін деп есептелінеді.
Екінші жағынан, сперматозоидтар өзара таласып бәсекелес алады, ондайда белгілі қасиеттері бар сперматозоидтар көбінесе жұмыртқа клетканы ұрықтандырады. Мұндай құбылыс селективті ұрықтану деп аталады. Бұл жағдай еркін шағылысуға (панмиксия) тежеу салады және өсімдіктер мен жануарлар эволюциясында оқшаулануға бейімделу механизмдерінін бірі болып табылады.
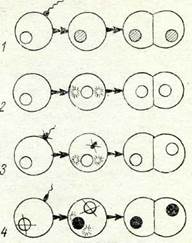 |
| 20-сурет Жыныстық жолмен кө-беюдің әр түрлі типтері: 1 —ңалыпты ұрыңтану; 2 —партено-генез; 3 —гиногенез; 4—андрогенез. |
5. ЖЫНЫСТЫ КӨБЕЮДІҢ ТҰРАҚСЫЗ ТИПТЕРІ
Жынысты көбеюдің тұрақсыз типтерінІң классификациясы. Жынысты кебеюдің тұрақсыз типтеріне жануарлар мен есімдіктерді партеногенездік, гипогенездік және андрогенездік көбеюлері жатады (20-су-рет).
Партеногенез — бұл ұрықтанбаған жұмыртқа клеткадан ұрықтың өсіп-жетілуі. / Табиғи партеногенез қүбылысы төменгі сатыдағы шаян тәрізділерге, коловраткаларға, жарғақ қа-наттыларға (араларға, сона-
ларға) және т. с. с. тән болады. Ол құбылыс құстарда (күрке тауықта) да кездеседі. Партеногенезді қолдан жасауға да болады, бұл үшін ұрықтанбаған жұмыртқа клеткаға әр түрлі агенттермен әсер етіп, оларды активтендіреді.
Партеногенезді денелік, яғни диплоидты және генеративті, яки гаплоидты деп беледі. Денелік партеногенез кезінде жұмыртқа клетка редукциялық жолмен бөлінбейді, бүл кезде гаплоидты екі ядро өзара косылып бірігеді де, хромосоманың диплоидты жиынтығын қалпына келтіреді (автокариогамия); сөйтіп ұрық тканінің клеткаларында хромосоманың диплоидты жиынтығы сақталады.
Генеративті партеногенезде ұрық гаплоидты жүмыртқа клеткадан өсіп-жетіледі, Мысалы, бал арасында (Аріз теііі-?ега) еркек аралар партеногенез жолымен ұрықтанбаған гаплоидты жүмыртқадан жетіледі.
Өсімдіктердегі партеногенезді көбінесе апомиксис деп атайды. Апомиксис өсімдіктер дүниесінде кең таралған және түқым қуалауды зерттеп білуде үлкен маңызы бар. Апомиксистік көбеюдің әсіресе көп таралған типі партеногенездік жолмен жұмыртқадан ұрықтың пайда болуы. Бүл кезде диплоидты апо миксис (мейозсыз) жиі кездеседі.
Эндосперм түзілу кезінде де, үрық түзілу кезінде де тұқым қуалаушы информация аналық организмнен беріледі. Бірқатар апомиксистерде толық қүнды тұқым түзілу үшін псевдогамия — тозаң түтігінің ұрық қалтасын активтендіруі қажет. Бүл кезде түтік тұқым қалтасына жеткеннен кейін, ондағы бір спермия ыдырап жойылады да, екінші спермия орталық ядромен қосы-лып бірігеді, сейтіп эндосперм ткаиінің түзілуіне ғана қатысады (Роіепііііа, КиЬиз т. с. с. туыстардың турлері). Мұндағы тұқым қуалаудың алдыңғы жағдайдан біршама айырмасы бар. Үрықта шешесінің белгілері ғана тұқым қуалайды, ал эндоспермде шешесінің де және әкесінің де белгілері тұқым қуалайды.
Гиногенез, Гиногенездік көбею партеногенезбен өте ұқсас болады. Гиногенездік кебеюдің партеногенезден айырмасы — мұнда жұмыртқа клеткасыныц өсіп-жетілуі үшін сперматозоидтар стимулятор қызметін атқарады (псевдогамия), бірақ бұл кезде үрықтану (кариогамия) болмайды; ұрық тех қана аналык ядродан өсіп-жетіледі (20-сурет, 3). Гиногенез жұмыр құрттар-да, тірі туатын балықта (Моіііепезіа іоптюза), күміс түсті та-бан балықта (Ріаіуроесііиз) болады._}Сол сияқты сарғалдақ (Раішпсиіиз аигісотиз), қоңырбас (Роа ргаіепзіз) т. б. бірқатар өсімдіктерде кездеседі.
Гиногенездік көбею құбылысының тұқым куалаушылықты зерттеуде үлкен маңызы бар, өйткені бұл кезде үрпақ түқым қуалаушы информацияны тек қана шешесінен алады.
Андрогенез. Гиногенезге қарама-қарсы қүбылыс андрогенез болып табылады. Андрогенез кезінде жүмыртқаның дамуы тек аталық ядро мен аналық цитоплазманың есебінен жүреді (20-сурет, 4). Ұрықтану сәтіне дейін аналық ядро қандай да болмасын бір себептен жойылып кетсе, андрогенез сонда ғана жүреді.
Егер жүмыртқа клеткасына бір сперматозоид түссе, хромосомалар жиынтығы гаплоидты жаңа дамып келе жаткан ұрықтың тіршілік қабілеті болмайды немесе ол нашар болады. Егер хромосомалар жиынтығы диплоидты қалыпқа келсе, онда андрогенді зиготалардың тіршілік қабілеті қалпына келеді. Жоғарыда айтылғандай, жануарларда полиспермия болған кезде жұмыртқа клеткаға бір мезгілде бірнеше сперматозоид енеді, бүл кезде аталық екі пронуклеус өзара қосылып бірігуі және диплоидты ядроның түзілуі мүмкін. Андрогендік организмнің дамуы тек тұт көбелегінен (ВотЬух гаогі) және паразит сонадан (НаЬгоЬгасоп ]ид1ап(ііз) байқалды. Андрогендік жолмен көбею ерекше құбылыс ретінде кейбір әсімдіктерде (темекі, жү-гері т. б.) кездеседі.
6. ТІРШІЛІК ЦИКЛІНДЕ ГАПЛОФАЗА МЕН ДИПЛОФАЗАНЫҢ








