 2015-06-28
2015-06-28 2689
2689Тіркесу тобын анықтау. Егер гендер хромосомада тізбектеліп орналасса, ал кроссинговердің жиілігі гендердің ара қа-шықтығын көрсетсе, онда геннің хромосомадағы орнын анық-тауға болады.
Геннің орнын, яғни оны локализациялау үшін ең алдымен ол геннің қандай хромосомада орналасқанын анықтау керек. Бір хромосомада орналасқан және өзара тіркесіп тұқым қуалайтын гендер тіркескен топ қүрады.
Анығында, әрбір түрдегі тіркескен топтар саны хромосома-лардың гаплоидты жиынтығына сәйкес келеді.
Қазіргі кезде тіркескен топтар генетикалық түрғыдан әсі-ресе жақсы зерттелген объектілерде анықталды. Барлық жағ-дайда тіркесулер тобы хромосомалардың гаплоидты санына то-лық сәйкес болатыны табылды. Мысалы жүгерідегі (2еа тауз) хромосомалардың гаплоидты саны мен тіркесулер тобы 10-ға тең, бүршақтікі (Різит заііуит) — 7, дрозофиланікі фгозорһііа теіапо^азіег) — 4, үй тышқанынікі (Миз тизсиіиз) — 20 және т. с. с.
Геннің белгілі бір тіркесулер тобына жататынын анықтау принципі белгілі тіркесулер тобында орналасқан басқа ген-дермен салыстырғанда осы геннің тұқым қуалау сипатын анық-тауға келеді. Мысалы бұршақтағы жеті тіркесулер тобыңың. (Ь, с, й және т. б.) әрқайсысында біреуден рецессивті гендер белгілі деп есептейік, рецессивті а гені тіркесулер тобының қайсысына жататынын анықтау қажет болсын дейік. Бұл ген тіркесулердің белгілі бір тобына ғана жататын болғандықтан, ол ген қалған алты тіркесулер тобынан тәуелсіз тұқым қуалауы тиіс. Мысалы, -. =, х % п ек* генотиптерінің особьтарын өзара шағылыстырғанда Ғ2-де ажырау мынадай болады: 9А-О--;ЗА-сісі:ЗааО:\аасі<і. Бірақ тіркескен жеті топтардың бі-реуінің генімен, мысалы с гені мен а геніиің тіркесуі толық Ас аС ■п емес дейік .^Х^ждіҒііаияағъі шағылысуы Ғ2 күтілген төрт класс А-С-, А-сс, ааС- және аасс береді, бірақ, олардың өзара қатынасы 9:3:3:1 болмайды да, ата-аналық кластар А-сс жә-не ааС —■ басым болады, ондай басымдық тіркеліс күшіне бай-ланысты не үлкен немесе шағын болады.
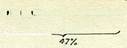 |
Бірақ кариотиптегі хромосомалардың нақты қайсы жұбы тиісті тіркестер тобына сәйкес > екенін генетикалық әдіспен анық- 1,1%3,5% тау мүмкін емес. Бүл үшін қосымша генетикалық зерттеулер жүргізу қажет. Соңғы уақытта тіркеулер тобын анықтау үшін дене 44-сурет. Хромосомадағы гендердің локализациялану схемасы. клеткаларын будаидастыру әдісін кроссинговер-пайдаланады (VIII бөлімді қаді цифрлар көрсвтеді. раңыз).
Генді локализациялау. Генге қатысты тіркесулер тобын анықтағаннан кейін, анализдін, келесі кезеңіне көшеді де, геннің тіркесулер тобындағы орнын табады. Генді локализациялау кроссинговер нәтижелерін есептеу жолымен іске асырылады. Кейде генді локализациялау үшін цитологиялық әдісті де пай-даланады, ол жөнінде алда баяндалады.
Хромосомадағы геннің локусын табу үшін шағылыстыруды мынадай жолмен жүргізеді: кроссинговерде анықталатын геи локусы қос қабат кроссинговер қүбылысындағыдай үшінші нүкте болуы тиіс. Хромосомдағы үш локуске маркировка салу гендердің орналасу ретін,ара қашықтығын анықтау үшін қажет.
Жоғарыда өзіміз қарастырғаи дрозофилаға қатысты мысал-да у және хю гендері арасындағы кроссинговер проценті 1,2-ге тең, ал ш және Ы гендері арасындағы кроссинговер проценті 3,5-ке тең. Бұл керсеткіштер бойынша у гені ш геннің оң жағында ма, сол жағында ма, әлі айтуға болмайды, сол сияқты хю ген-нің, Ы генімен салыстырғандағы орналасу жағдайы жөнінде де ештеңе айтуға болмайды. Гендердің үшінші жүбы у және Ы гендері аралығындағы айқасу процентін анықтағаннан кейін ғана (біздің жағдайда 4,7%) т гені, у және Ы гендері аралығына орналасқан болуы тиіс деген қорытындыға келуге болады. Тіркесу тобында ген белгілі бір орын алатындықтан, бүл хромо-сомадағы гёндердің орналасу тәртібіи анықтауға және хромосомалардың генетикалық картасын жасауға мүмкіндік береді.
5. ГЕНЕТИКАЛЫҚ КАРТАЛАР
Генетикалық карта жөнінде түсінік. Белгілі бір тіркесулер тобына кіретін гендердің салыстырмалы түрде орналасу схема-сын хромосомалардың генетикалыщ картасы деп атайды. Гене-тикалық көзқарас түрғысынаи алғанда әсіресе көбірек зерттел-ген кейбір: дрозофила х жүгері, томат, тышқан, нейроспора, ішек таяқшасына ғана әзірше ондай карталар жасалды.
Генетикалық карталар гомологиялық хромосомалардың әр жүбы үшін жасалады. Тіркесулер тобын нәмірлейді.

45-сурет. Жүгері хромосомаларының генетикалық картасы:
1 — X — тіркесу топтары: цифрлар гендердің ара ңашыңтығын көрсетеді (тіркесу өлшемімен). Центромерлер дөцгелекпен көрсетілген. Гендердіц атаулары:I: зг—ала жапыраңтар; дав — гаметофит факторы; тзп — аталың стерильділігі;івг— түңымы бар шашаңтар; Р — боялған перикарп; гі — зиготалың леталь; аз — асинапсис; һт — гельминтоспориозға шыдамдылың; ңысңарған буын аралың; шашаң негізі; /і— жіңішке сызыңшалы жапыраңтар; ащ — аталығы бар сабаңтар; Кп — бунаңталған жапыраңтар; жасыл жолаңты жапыраңтар; Тзс — тұңымы бар шаіпаңтар; Ьт2 — жапырақ ортасындағы ңоңыр түсті өзек.II: -и)83—аң түсті орам; аі —аңшыл жаиырадтар; лигульсіздік; §Һ —жылтыраған жапырақтар; В —антоциаи бояуларын күшейтуші; зһ —«шелка» жоңтығы; І\х — күңгірт эндосперм; І8\ — түңымы бар шашақ; «4— сарғыш жасыл өскіндер; Сһ— шоколад түстес перикарп.III: сг\ — бүралган жапыраңтар; й\ — ергешейлілік; гі — тамырдың болмауы; Ь§3—лигулсіздік; В бүжырлы жапыраңтар; із, — түңымы бар жапырақтар; Ъа\ —үрпаңсыз сабаңтар; пщ — ерғежейлілік; йі— қоңыр пертткарп; в/г2— бүріскен эндосперм; еі — оіоланган эндосперм; да\ — гаметофит факторы.IV: йе% — жетілмеген әндосперм; Са< — гаметофит факторы; Тв түңымы бар шашаң; Зрі — үсаң тозаң; 8мі— ңантты эндосперм; б,е№ — жетілмеген эндосперм; 2Ь6—көлденең жолаңты жапыраңтар; жарғаңты;;2—«жапондьщ» ала жолаңтың; §Һ — жылтыр жапыраңтар.V: жылтыр жапыраңтар; а2— өсімдіктің антоциан бояуы; Ьт\ — қоңыр түсті ортаңғы өзек. Ы\ — нәзік өндосперм; г;3— сарғыш жасыл түсті өскіндер; Ьг?і— ңысңа всімдік; рг — ңызыл алейрон; сары жолаңтың; г>а— сарғыш жасыл түсті ескіндер.VI: ро] — көптеғен митоздар; у\ — сары эндосиерм; рд^ — ашың жасыл өскіндер; РІ —•ңара ңошңыл өсімдік; Вһ — таңдаңтьт алейрон; ат, — аналың аузының ңызгылт бояуы және «шелка», ру —үсаң өсімдік.VII: Нз —түк басңан орам; іп —алейрон бояуын күшейтуші; Vх— сарғыш жасылтүсті ескіндер; га^— тармадталған масаң; £һ — жылтыраң жапырак.тар; Трі— еягер-ген гүл шоғары; 8?—кертілгеп жапыраңтар; і?—ала түстілік; Вп— ңоңыр алейрон; Ъй —«шелкасыз» тармаңталған сабаң.VIII: «,6—сары жасыл өскіндер; тве, — атальщтың стерильділігі; іі—«жопандың» жолаңтың.Картаны жасау үшін көптеген гендердің тұқым куалау зақдылықтарын зерттеп білу қажет. Мысалы, дрозофиланын төрт тіркесулер тобына жинақталған 500-ден астам жүгерінің он тіркесулер тобына жиналған 400-ден астам гендері зерттелген.
Генетикалық карталар жасағанда тіркесулер тобы, гендер-дің толық немесе қысқартылған аттары, хромосоманың ноль ретінде қабылданған бір шетінен бастап %-пен көрсетілген қа-, шықтығы керсетіледі; центромераның орны белгіленеді.
45-суретте қысқартылған схема түрінде келтірілген жүгері хромосомаларыиың карталары көптеген ғалымдардың жүйелі еңбектерінің нәтижесі болып табылады. Генетикалық картаның ұзындығы хромосоманың мелшеріне байланысты.
Генетикалық карталар жасау гендері картаға түсірілген белгілердің түқым қуалау сипатын болжап айтуға мүмкіндік береді, ал ол селекциялық жүмыстарда шағылыстыру үшін ата-аналар жұбын таңдап алу да жеңілдік келтіреді.
Хромосомалардың генетикалық карталарын қараған кезде мынадай сүрақ тууы мүмкін: геннің алатын орны 62 немесе 107% болса, жүгерінің тіркесулер тобындағы геннің локусы қалай анықталады? Ал дигетерозигота түзетін кроссоверлі га-металар тіпті 50%-ке тең бола алмайды, өйткені гаметалардың ата-аналар мен гендердің жаңа үйлесімдерінің мұндай қаты-наста болуы тек тәуелсіз тұқым қуалау кезінде ғана байқала-ды. Демек, бір хромосома шегіндегі ең шеткі екі нүктенің ара қашықтығы 50% бола алмайды. Мүндай сәйкессіздік сияқты болып көрінуді былай түсіндіруге болады: гендер локализация кысқа және ретімен алынатын учаскелердің кроссинговерле-рін есептеу арқылы іске асырылады, ал барлық учаскелер үшін анықталған кроссинговер мөлшерінің жиынтығы қартаға түсіріледі. Сондықтан генетикалық картаның жалпы үзындығы эксперимент нәтижесінде алынған хромосоманың қарама-қарсы шеттерінде орналасқан гендер арасындағы кроссинговер шама-сынан көп үлкен болуы мүмкін.
Микроорганизмдердің генетикалық карталары. Көп клетка-лы организмдерде гендер рекомбинациясы реципрокты болады. Ал микроорганизмдерде ол бір жақты болуы мүмкін. Мысалы, біркатар бактерияларда, атап айтқанда ішек таяқшасында (Езсһегһіа соіі) генетикалық информацияның алмасуы клетка-лар конъюгациясы кезінде іске асады (15-тарауды қараңыз). Бактерияның тұйық сақина тәріздес жалғыз хромосомасы конъюгация кезінде үнемі белгілі бір нүктеден үзіледі де бір клеткадан екінші клеткаға ауысады.
Берілген хромосома учаскесінің үзындығы конъюгация үзақтығына байланысты болады. Хромосомада орналасқан гендер
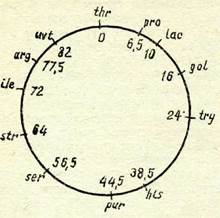
46-сурет. Езсһегісһіа соіі генети-калық картасы.
Геидердің ара ңашыңтығы минут арқылы берілген. Гендердің белгісі: іһг, рго, Іту, һгз, риг, зег, ііе, аг£ —-троенинді, пролинді, трифтофанды, гистидинді, пуринді, серинді, изо~ лейцинді, аргининді ңажет етуі; Іаз, §аі — лактоза мен галактозаны ашытуға ңабілетсіздігі, віг, иог —■ стрептомицинге, ультра-кулгінге тұраңты-отың;
реті тұрақты болады. Осыған байланысты мұндай сақиналы картада гендердің ара қашықтығы кроссинговер процентімен өлшенбейді, коньюгация ұзақтығын керсететін минутпен өлше-неді (46-сурет).
6. ТЕТРАДАЛЫҚ АНАЛИЗ КЕЗІНДЕ КРОССИНГОВЕРДІ ЕСЕПТЕУ
Жоғары сатыдағы организмдерде мейоздың профазасында өткен кроссинговерді кроссоверлі особыардың — рекомбинанттардың жиілігі бойынша біледі, өйткені олардың пайда болуы кроссоверлі және кроссоверлі емес гаметалардың' қатынасын көрсетеді деп есептеледі.
Рекомбинантты зиготалардың кроссоверлі гаметаларға сәйкес келетіндігін тікелей дәлелдеу үшін, кроссинговердің нәтижелерін мейоздың гаплоидты өнімі бойынша тікелей анықтау қажет. Бұл кезде гендер өздерінің әсерін гаплофазада көрсетуі тиіс. Осындай зерттеу іске асқан объекті мысалы, зең саңырау-құлағы (Меигозрога сгазза) болды. Бұл саңырауқұлақта тірші-лік циклінің негізгі бөлігі гаплофазада өтеді, ал оның диплоидтық фазасы өте қысқа болады.
Ұрықтанғаннан кейін көп ұзамай, зиготада мейоздық бөліну басталады да, гаплоидты сгюралары бар қалташалар—аска түзілуіне алып келеді. Бөліну кезінде ұрық осі қалташаның көл-денең осіне сәйкес келеді. Сондықтан мейоз өнімі—споралар—-қалтаның ішІнде тізбектеле орналасқан. Мейозда пісіп-жетілудіқ қалыпты екі бәлінуі, бүдан кейін бір митоздық бөліну болады да, осының нәтижесінде әр қалташада 8 аскоспора түзіледі.
Мейоз өнімі бойынша кроссинговер нәтижесін тікелей анықтауға нейроспора мүмкіндік беретіыдіктен, бұл жағдайда ажырау сипатын анықтау, кроссинговермен ажырау мейозда іске асатындығынын, тікелей дәлелі болып табылады. Бүл әдіс жоғарыда баяндалған тетрадалық аиализдің бір түріне жатады, бірақ мұның тіркескен гендерге қатысы бар.

47-сурет. Нейроспорада аскоспоралар түзу кезіндегі гомологиялық хромосомалар қасиетінің әр түрлі типті схемасы: 1 — кроссинговер байңалмайды; 2 — кроссинговер байқалады.Моногибридті шағылыстыру жағдайында гаплоидты өнім-дер (споралар) бойынша ажырау 1 А: 1 а қатынасында болады деп күтіледі. 47-суретте аскоспоралардың реңін анықтайтын ал-лельдердің бір жұбы бойынша (А және а) ажырау келтірілді. Аскадағы 8 спораның 4-нің боялғаны (Л), 4-нің боялмағаны (а) көрінеді, яғни ажырау 1: 1 қатынасында болатыны байқалады. Ген мен центромера аралығында кроссинговер жоқ кезде спо-ралардың қалтада орналасу тәртібі мынадай: АЛАЛаааа (47-сурет, 1). Егёр аскоспоралардың орналасу тэртібі, мысалы, ААааААаа болып өзгерсе, онда ол а локусы мен центромера арасында өткен айқасу болғандығын көрсетеді (47-сурет, 2).
Споралардың орналасуы бірінші және екінші мейоздық бө-лінулер кезіндегі хромосомалардың таралуынан тәуелді болады. А және а аллельдері қалташықтағы споралар бойынша басқаша да ретпен орналасуы мүмкін.
Біз қарастырып отырған жағдайда айқасу осы геннің локусы мен центромера арасындағы учаскеде болады, а гені центромерадан қашықтаған сайын, айқасу мүмкіндігі арта береді, демек кроссоверлі аскалар саны да көбейеді. Егер айқасу хромосоманың дистальді шеті.мен а гені арасында болса, онда аскоспоралардың кроссоверлі орналасуы табылмайды (47-су-рет, 1). Ген мен центромера арасындағы кроссинговер кезінде ас-кадағы споралардың орналасу тәртібінің өзгеруі тек ол төрт жіпше стадиясында яғни хроматидтер арасында өтсе ғаиа мүм-кін болады. Егер рекомбинация әр хромосома әлі қосарланбаған кезде өтсе, онда аскадағы споралар тәртібі өзгермес едь Демек, споралардың орналасу тәртібінің өзгеруі бұл жағдайда кроссинговердің сіңлілі емес хроматидтер арасында, яғни төрт жіпшелер стадиясында жүргендігінің дәлелі болып табылады.
Сондықтан кроссинговердің механизмі және оның генетика-лық с&лддры жөиіаде айтқак кезд,е, ондшіату үщін ғана сжы тұтас хромосомалар арасындағы алмасу деп түсіндіреді, ал шындығында алмасу хроматидтер арасында жүреді. Нейроспо-раның аталған ерекшеліктері аллельдердің бір жүбы бойынша ажырауды ескере отырып, геннің хромосомадағы орнын анықтауға мүмкіндік береді; тетрадалық анализ жасауға болмай-тындықтан диплоидты организмдерде бүл мүмкін емес.
Сонымен тетрадалық анализ мендельдік ажыраудың, сол сиякты кроссинговердің де мейоздың' заңдылықтарына негізделгенін дәлелдейді.
7. КРОССИИГОВЕРДІҢ ЦИТОЛОГИЯЛЫҚ ДӘЛЕЛДЕРІ
Кроссинговер құбылысын генетикалық әдістермен анықта-ғаннан кейін, гендер рекомбинацияларымен қоса жүретін гомологиялық хромосомалар учаскелерінің алмасатынына тікелей дәлел алу қажет еді. Мейоздың профазасында байқалатын хи-азма бейнесі қүбылыстың тек жанама дәлелі ғана бола алады, өткен алмасуды тікелей бақылау мүмкін емес, ейткені учаскелерімен алмасушы гомологиялық хромосомалардың, әдетте, мөлшері де, пішіні де тіпті бірдей болады.
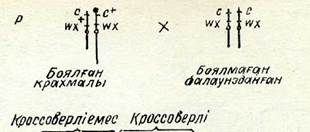
Г. Крейтон мен Б. Мак-Клинток гомологиялық хромосома-ларында морфологиялық айырмасы бар жүгерінің формасын алды:— біреуі қалыптағыдай, екіншісінің бір иығының шеті жуандау, екінші иығы үзарған. Хромосома жүптары құрылысындағы бүл ерекшеліктер цитологиялық зерттеулер кезінде оңай табылады (48-сурет).
Тәжірибеде қалыпты хромосомада рецессивті геи с (боялмаған эндосперм) және доминантты ген шх+ (крахмалды эндосперм), ал өзгерген хромосомада доминантты ген с+ (боялған эндосперм) және рецессивті ген шх (балауыз тәрізді эндосперм)
бар екені анықталды. Дигетерозигота - морфологиялық қалыпты хромосомасымен қоса таңбаланған рецессивті гендері с және шх бар линиялармен шағылыстырды. Ұрпағында кроссоверлі емес те және кроссоверлі де дәндер алынды. Оларды цитоловиялық зерттегенде кроссоверлі дәндерде алмасқан учаскелері бар хромосомалардың үнемі болатындығы табылды: ұзындығы қалыптағыдай, бірақ жуандауымен (утолщением) немесе үзынырақ жуан емес (без утолщения).
 |
Сонымен бір мезгілде цитологиялық та, генетикалық та гендердің рекомбинациясы мейоздың профазасында гомологиялық. хромосомалар учаскелерінің алмасуымен қоса жүретіндігі анықталды.
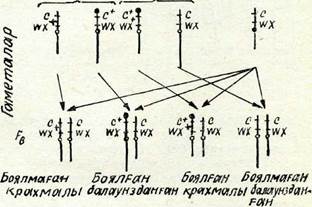 |
48-сурет. Жүгеріде болатын кроссинговердің цитоло-гиялық дәлелі:
с +— боялған әндосперм; с— боялмаған эндосперм; адх + крахмалды әндоссперм; гох— балауыз тәрізді эндо-сперм.
8. ХРОМОСОМАЛАРДЫҢ ГЕНЕТИКАЛЫҚ








