 2015-06-26
2015-06-26 1016
1016ЭЭГ – результат алгебраического суммации постсинаптических потенциалов корковых нейронов.
Потенциалы действия нейрона вносят незначительный вклад в процесс формирования ЭЭГ.
Наиболее эффективно суммация происходит на мембране нейрона при одновременном синхронном поступлении возбуждения по многим синапсам. При этом возникают высоко амплитудные волны ЭЭГ.
Альфа ритм: причины возникновения синхронных синоптических изменений на мембране корковых нейронов. Является циклические таламокортикальные взаимодействия. Возбуждение рецепторов попадает в кору головного мозга порциями через таламические нейроны, играющие роль ритмо-водителей – пейсмекеров.
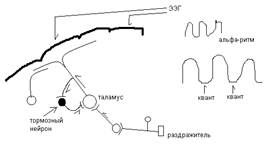
Один из механизмов ритмической активации таламических нейронов, который разбивают на кванты поступающее воздействие считают воздействие. Самоторможение нейронов, создающее периодические колебания возбудимости нейронов с помощью тормозных нейронов (ТН). Кроме того на таламус влияют импульсы из ретикулярной фармации ствола мозга.
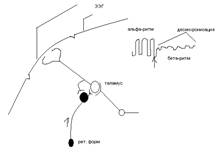
В ретикулярной формации имеются тормозные нейроны, которые угнетают активность тормозных нейронов таламических ядер и в этом случае возбуждение начинат поступать к корковым нейронам не порциями а сплошным потоком – формируется стабильный электрический потенциал, уменьшается амплитуда волн ЭЭГ, увеличивается их частота – наблюдается реакция активации, десинхронизации ритмов ЭЭГ.
ЭЭГ применяют в клинике для:
- Для общей оценки деятельности мозга – человек умирает, пульс не прощупывается дыхание отсутствует. ЭЭГ даст ответ жив мозг или нет (следует проводить реанимацию или нет).
- Применяют ЭЭГ при оценке глубины наркоза с целью предупреждения передозировки.
- Для диагностики многих состояний – травма мозга, инсульт, опухоли и т. д.. Обычно поврежденный участок мозга теряет или снижает электрическую активность или нарушается нормальная ритмика.
МЕТОД ВЫЗВАННЫХ ПОТЕНЦИАЛОВ (ВП)
Используется для изучения функций коры головного мозга. ВП представляет собой стабильный комплекс волн на ЭЭГ, который возникает при однократном нанесении раздражения на периферический нерв, однородную группу рецепторов.
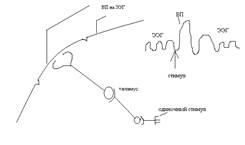
Изобразим ВП в большем масштабе:
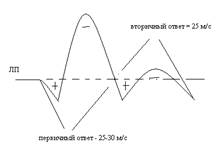
ВП наступает не сразу а спустя латентный период (10-15 м/м). ЛП зависит от времени проведения возбуждения с периферии в кору головного мозга. Далее возникает ВП, который представляет собой производное от двух потенциалов: начальный компонент (первичную позитивную волну + и первичную негативную волну -, это первичный ответ); вторичный ответ (ВО – вторичная позитивная и негативная волна).
Отличие ПО от ВО принципиально. По регистрируется в строго определенной части коры в зоне представительства определенного нерва (проекционной зоне). ВО регистрируется во всех участках коры, генерализованый ответ. В проекционной зоне ВО подстраивается к ПО и выглядит как его продолжения, но это самостоятельный тип электрической реакции.
ПО выражает процесс обработки, прихода возбуждения в корковый проекционный пункт по таламокортикальному тракту.
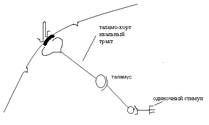
ВО возникает в следствии иррадиации (распространения) возбуждения через ядра ретикулярной формации (РФ), которые по ретикуло-кортикальным путям поступают во все пункты коры.
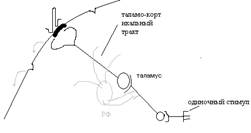
С помощью ВП можно:
1. изучать локализацию проекционных зон в коре (картирование мозга). Затылочная доля возбуждение от рецепторов сетчатки; Слуховое раздражение ВП в области височной кости.
2. по ВП можно изучать закономерности проведения возбуждения по структурам мозга (например при поражении РФ искажается или пропадает вторичный ответ.
3. можно изучать действие лекарственных веществ на кору и ГМ, препараты могут по разному влиять на параметры ВП (он может увеличивается или уменьшатся по амплитуде и протяженности как весь так и в отдельных компонентах)
СТЕРЕОТАКСИЧЕСКИЙ МЕТОД
Стереотаксис (движение чего то в объеме – перевод). Этот метод позволяет проводить манипуляции с подкорковыми структурами, подводя к ним инструменты или электроды в слепую.
В подкорковые ядра вводят инструменты или электроды с целью:
1. Регистрации электрической активности ядра
2. Для стимуляции ядер (электрической или химической)
3. Коагуляции – разрушений ядер
Размеры и форма черепа у отдельных видов и разных людей почти одинаковы – разброс незначителен. С точки зрения геометрии мозга у всех особей подкорковые структуры локализованы одинаково. Возникла идея описать каждое ядро относительно стандартных координат – стереотаксических плоскостей.
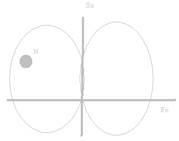
Посмотрим на мозг кошки сверху. Проведем гипотетическую линию по стреловидному шву черепа, через нее также можно повести и плоскость – нулевая сагиттальная плоскость (S0). Проведем еще одну плоскость (F0), для чего используют стабильный ориентир (нижний край глазницы + наружный слуховой проход. Через эти две точки мы можем провести базовую линию (собственно нулевая фронтальная плоскость проводится перпендикулярно к базовой линии ч/з центр слуховых проходов.
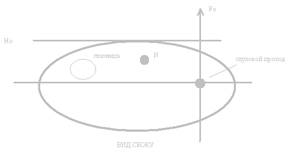
Для точного введения нужна третья координата, которая определяет глубину погружения электрода. Горизонтальная плоскость (Но).
Относительно этих трех координат анатомы описали расположение всех подкорковых образований (в мм) и объединили их в специальные атласы «Атласы стереотаксических карт мозга» (для человека, кошки, мыши и т. д.). В настоящее время в дополнении к стереотаксической методики применяют дополнительные критерии правильности введения электродов.








