 2015-07-04
2015-07-04 3841
3841При изучении сетчатки необходимо выяснить следующие проблемы: во-первых, каким образом палочки и колбочки преобразуют поглощаемый ими свет в электрические и химические сигналы; во - вторых, как последующие клетки двух других слоев - биполяры, горизонтальные, амакриновые и ганглиозные - интерпретируют эту информацию.
Молекулярная физиология фоторецепции
Сетчатка по своему строению и происхождению представляет собой нервный центр, в котором происходит первичная обработка зрительных сигналов, преобразование их в нервные импульсы, передающиеся в головной мозг.
Она имеет сложную многослойную структуру. Здесь расположены два вида вторично-чувствующих, различных по своему функциональному значению фоторецепторов и несколько видов нервных клеток (рис.21.).
Возбуждение фоторецепторов активирует первую нервную клетку сетчатки - биполярный нейрон. Возбуждение биполярных нейронов активирует ганглиозные клетки сетчатки, передающие свои импульсы в подкорковые зрительные центры. В процессах передачи и переработки информации в сетчатке участвуют также горизонтальные и амакриновые клетки. Все перечисленные нейроны сетчатки с их отростками образуют нервный аппарат глаза, который не только передает информацию в зрительные центры мозга, но и участвует в ее анализе и переработке. Поэтому сетчатку называют «частью мозга, вынесенной на периферию».
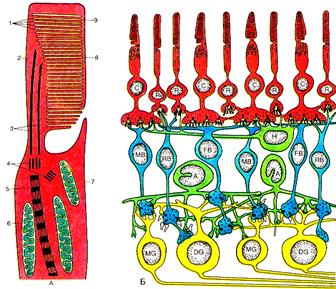
Рис.21. Строение фоторецептора (А) и сетчатки глаза (Б) С-колбочки; R-палочки; Н-горизонтальная клетка; МВ,RВ; FD- биполярные клетки; А-амакриновые клетки; MG DG –ганглиозные клетки; 1-фрагмент диска; 2- наружный сегмент; 3- внутренний сегмент; 4- митохондрии внутреннего сегмента; 5- корешок реснички; 6- центриоли; 7-микротрубочки реснички; 8 – наружная плазматическая мембрана; 9- диски наружного сегмента. (воспроизводится по В.М.Покровскому,1998)
Внутриклеточные регистрации от колбочек и палочек животных показали, что в темноте вдоль фоторецептора течет темновой ток, выходящий из внутреннего сегмента и входящий в наружный сегмент. Освещение приводит к блокаде этого тока. Рецепторный потенциал модулирует выделение медиатора (глутамата) в синапсе фоторецептора.
Было показано, что в темноте фоторецептор непрерывно выделяет медиатор, который действует деполяризующим образом на мембраны постсинаптических отростков горизонтальных и биполярных клеток. На рис. 22 показаны направления ионных токов, текущих через плазматическую мембрану фоторецептора в темноте. В темноте плазматическая мембрана поляризована и на ней регистрируется разность потенциалов 35-45 мВ, обусловленная избытком отрицательных зарядов на внутренней поверхности и положительных на внешней. Градиенты концентрации Na+ и К+ поддерживаются на плазматической мембране палочки активной работой натрий-калиевого насоса, локализованного в мембране внутреннего сегмента.
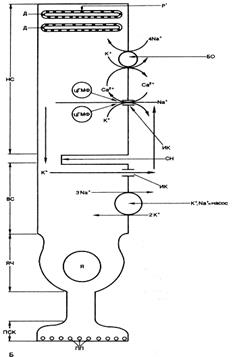 Рис. 22. Фотохимические процессы в палочковом аппарате сетчатки. Ионные токи через наружную мембрану палочки в темноте.Д - диск; СН - соединительная ножка; ЯЧ - ядерная часть; ПП - пресинаптические пузырьки; НС -наружный сегмент; ВС - внутренний сегмент; ПСК - пресинаптический комплекс; Я - ядро. (воспроизводится по В.М.Покровскому, 1998).
Рис. 22. Фотохимические процессы в палочковом аппарате сетчатки. Ионные токи через наружную мембрану палочки в темноте.Д - диск; СН - соединительная ножка; ЯЧ - ядерная часть; ПП - пресинаптические пузырьки; НС -наружный сегмент; ВС - внутренний сегмент; ПСК - пресинаптический комплекс; Я - ядро. (воспроизводится по В.М.Покровскому, 1998).
Уменьшение или прекращение входа внутрь наружного сегмента Na+ приводит к гиперполяризации клеточной мембраны, т.е. к возникновению на ней рецепторного потенциала.
Палочки и колбочки обладают уникальной среди всех рецепторов электрической активностью, их рецепторные потенциалы при действии света - гиперполяризующие, потенциалы действия под их влиянием не возникают. Палочки сетчатки имеют наибольшую чувствительность к длинам волн около 500 нм. Процесс поглощения света осуществляется в них фоточувствительными молекулами родопсина, которые в больших количествах(около 60 тыс. на диск) включены в структуру мембран дисков, размещенных стопками в наружных сегментах фоторецепторных клеток. Молекула родопсина представляет собой семисегментный трансмембранный(7ТМС) рецептор, ассоциированный с G белком. При поглощении света молекулой зрительного пигмента - родопсина в ней происходит мгновенная изомеризация ее хромофорной группы: 11-цис-ретиналь превращается в транс–ретиналь (Рис.23, 24). Вслед за фотоизомеризацией ретиналя происходят пространственные изменения в белковой части молекулы: она обесцвечивается и переходит в состояние метородопсина II. В результате этого молекула зрительного пигмента приобретает способность к взаимодействию с другим примембранным ассоциированным G белком – трансдуцином (Т) (– гуанозин трифосфат (ГТФ) - связывающим белком). Активированный в фоторецепторных клетках G белок (трансдуцин) диссоциирует с образованием Gα- субъединицы, которая связывается с ГТФ(в комплексе с метародопсином трансдуцин переходит в активное состояние и обменивает связанный с ним в темноте гуанозиндифосфат (ГДФ) на ГТФ). Трансфдуцин + ГТФ, активируют молекулу другого примембранного белка - фермента фосфодиэстеразы (ФДЭ). Активированная ФДЭ разрушает несколько тысяч молекул цГМФ.
В результате падает концентрация цГМФ в цитоплазме наружного сегмента рецептора. Это приводит к закрытию ионных каналов в плазматической мембране наружного сегмента, которые были открыты в темноте и через которые внутрь клетки входили Na+ и Ca2+. Ионные каналы закрываются вследствие того, что падает концентрация цГМФ, которая держала каналы открытыми. В настоящее время выяснено, что поры в рецепторе открываются благодаря цГМФ - циклическому гуанозинмонофосфату.
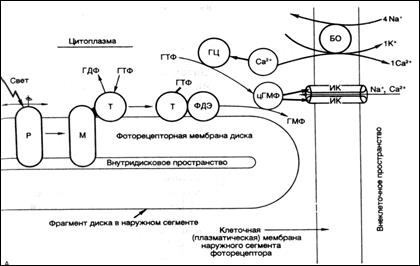
Рис. 24. Фотохимические процессы в палочковом аппарате сетчатки. Фрагмент фоторецепторного диска. Р-молекула родопсина; М-метародопстн II; БО-белок-обменник; ИК-ионный канал; Т-трансдуцин; ФДЭ-фосфодиэстераза; ГЦ-гуанилатциклаза; цГМФ-циклический гуанозинтрифосфат; ГМФ-гуанозинмонофосфат; ГДФ-гуанозиндифосфат; ГТФ - гуанозинтрифосфат (воспроизводится по В.М.Покровскому, 1998).
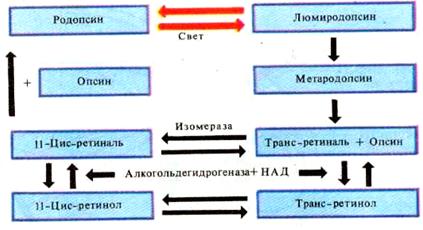
Рис. 25. Схема образования и обесцвечивания родопсина
ТРАНСФОРМАЦИЯ СВЕТОВОЙ ЭНЕРГИИ В НЕРВНОЕ ВОЗБУЖДЕНИЕ
 Квант света
Квант света
Активация родопсина: свет, поглощаемый родопсином, инициирует ответ в каскаде цГМФ
 |
Каталитическая активация G-белка родопсином
 |
Активация циклической фосфодиэстеразы
 |
Гидролиз цГМФ активированной цГМФ-фосфодиэстеразой
Закрытие цГМФ зависимых Na+ каналов
Гиперполяризация фоторецепторов сетчатки
Передача сигнала на биполярные клетки: изменение характера секреции медиатора в синапсе между внутренним сегментом рецепторной клетки и дендритом биполярной клетки
Механизм восстановления исходного темного состояния фоторецептора связан с повышением концентрации цГМФ (в темновую фазу с участием алкагольдегидрогеназы + НАДФ). Снижение проницаемости натриевых каналов при действии света сопровождается также снижением поступления в клетку ионов Са2+, уменьшение его концентрации в клетке ведет к повышению активности фермента гуанилатциклазы, увеличивающей скорость образования цГМФ из ГМФ. Уменьшение концентрации ионов Са2+ также приводит к снижению активности фермента фосфодиэстеразы, разрушающей цГМФ. В результате всего этого концентрация цГМФ внутри клетки повышается и цГМФ вновь связывается с ионным каналом плазматической мембраны, открывая его. Через открытый канал внутрь клетки вновь начинают входить Na+ и Ca2+, деполяризуя мембрану рецептора и переводя его в «темновое» состояние. Из пресинаптического окончания деполяризованного рецептора вновь ускоряется выход медиатора. Предполагается, что от уровня кальция существенно зависит скорость восстановления чувствительности(адаптации) фоторецепторов как в условиях высокой, так и низкой освещенности.
Т.о., поглощение света, молекулами фотопигмента приводит к снижению проницаемости для Nа+, что сопровождается гиперполяризацией, т.е. возникновением рецепторного потенциала. Гиперполяризационный рецепторный потенциал, возникший на мембране наружного сегмента, распространяется затем вдоль клетки до ее пресинаптического окончания и приводит к уменьшению скорости выделения медиатора - глутамата. Кроме глутамата нейроны сетчатки могут синтезировать и другие нейромедиаторы, такие как ацетилхолин, дофамин, глицин и ГАМК. На уровне рецепторов происходит торможение, и сигнал колбочки перестает отражать число поглощенных фотонов, а несет информацию о распределении и интенсивности света, падающего на сетчатку в окрестностях рецептора.
Механизмы восприятия и преобразования световых сигналов колбочками схожи с таковыми для палочек. Основные отличия реагирования колбочек на световые воздействия связаны с их более низкой (~в 70 раз) фоточувствительностью по сравнению с палочками.

