 2015-07-03
2015-07-03 1461
1461Существуют ли в стебле промежуточные двигатели восходящего водного тока? Дальний транспорт воды по стеблю осуществляется по ксилеме. Ксилема специально приспособлена к выполнению функции водопроведения. Проводящие элементы этой ткани имеют одревесневшие клеточные стенки и лишены протопластов. Сосудистые растения имеют два типа проводящих элементов ксилемы – членики сосудов (у покрытосеменных) и эволюционно более примитивные трахеиды (у голосеменных и папоротникообразных). Концы трахеид заострены. Они прикасаются боковыми стенками или заостренными концами. Контакт между ними осуществляется за счет окаймленных пор (расположенных строго друг против друга), перегородки которых состоят из первичной клеточной оболочки и срединной пластинки. Вследствие этого, вода должна просачиваться через указанный барьер. Сосуды представляют более совершенную водопроводящую систему. Гидравлическое сопротивление их меньше, чем у трахеид, поскольку диаметр больше и наличие перфораций на концах члеников сосудов делает движение воды более свободным по сравнению с её движением через окаймленные поры.
Большинство исследователей до сих пор полагают, что дальний транспорт воды по стеблю (ксилеме) представляет собой пассивное движение по непрерывному, плавному градиенту водного потенциала в системе почва-растение-атмосфера при участии двух концевых двигателей – нижнего (корневое давление) и верхнего (присасывающее действие транспирации). Движению воды по проводящей системе способствует взаимодействие молекул воды друг с другом и внутренней поверхностью ксилемы, то есть силы сцепления – когезия и адгезия. За когезию отвечают водородные связи между молекулами воды, а адгезию – водородные связи, образующиеся между молекулами воды и полярными группами соединений, входящих в состав ксилемы. Силы когезии обеспечивают непрерывность и высокую прочность водных нитей в условиях интенсивной транспирации, когда нити растягиваются и испытывают высокую нагрузку на разрыв.
Эти авторы полагают, что никаких специальных промежуточных двигателей водного тока в стебле нет. Ложе водного тока при движении воды по стеблю остается совершенно пассивным, как пассивны водопроводные трубы. При этом ссылаются на исследования Е.Ф. Вотчала (1897), который установил, что по положенному горизонтально отрезку древесного ствола длиной в несколько метров вода с одинаковой скоростью перемещается как от нижнего конца к верхнему, так и наоборот. Следовательно, он считал, что в древесине отсутствуют какие-либо клапаны, не пропускающие воду вниз, а без таких клапанов промежуточные двигатели якобы не могли работать. В других исследованиях показано, что введенные в сосуды ядовитые растворы (пикриновая кислота) поднимаются вверх по стеблю, вплоть до листьев, не отравляя на своем пути живые клетки. Беспрепятственное передвижение воды по стеблю наблюдали и при нагревании или замораживании участка стебля. Однако через некоторое время убитые участки стеблей переставали проводить воду, и в сосудах появлялся воздух.
Автоволновая динамика транспорта воды по стеблю. В работах А.А. Зялалова вместо плавного акропетального градиента водного потенциала ксилемного сока вдоль стебля установлены локальные градиенты в каждом отдельном междоузлии. Применив специальный датчик, он обнаружил локальные колебания тургарного давления паренхимных клеток, а также микроколебания диаметра стебля. Важно, что фазы микроколебаний по междоузлиям не совпадают. Каждое междоузлие ведет себя автономно: когда одно междоузлие сжимается, другое расслабляется и наоборот, то есть изменение диаметра всего стебля в целом подобно бегущей волне и напоминает перистальтику кишечника. Благодаря таким колебаниям ложе водного тока не остается пассивным: оно то сужается, то расширяется. Промежуточными двигателями восходящего водного тока могут быть перистальтические микроколебания объёмов паренхимных клеток стебля, изменяющие просветы сосудов ксилемы и тем самым способствующие проталкиванию воды вверх. В пользу такого предположения свидетельствует существование стеблевого давления (А.П. Петров).
Стеблевое давление. А.П. Петров обнаружил стеблевое давление в черенках стеблей клена, осины, березы при погружении их нижним концом в воду. В этих условиях они, не имея корней, выделяли экссудат. Это происходило за счет того, что живые клетки паренхимной обкладки проводящих пучков не только поглощали воду, но и нагнетали её в сосуды и проталкивали вверх по стеблю. По аналогии с корневым, стеблевое давление оказалось величиной аддитивной, то есть зависело от длины черенка, а иными словами, от количества паренхимных клеток обкладки. Причем, экссудация черенка (по аналогии с корневой экссудацией) имела автоколебательный характер (рис. 13). Эти данные доказывают существование промежуточных двигателей восходящего водного тока в стебле.
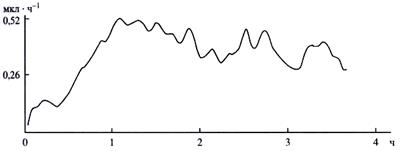 |
Рис. 13. Автоколебания экссудации черенка Acer platanoides.
Перистальтическая волна микропульсаций паренхимных клеток как метаболический двигатель восходящего водного тока. Как известно, индийский физиолог Д.Ч. Бос был противником представления о подъёме соков вверх лишь в результате «подтягивания сверху» и «подталкивания снизу», осуществляемыми листьями и корнями, или же благодаря только осмотическим процессам, которые «чрезвычайно медлительны», поэтому не могут обеспечить подъём соков на большую высоту. Он использовал специальный зонд, регистрирующий электрические сигналы в стебле. Д.Ч. Бос обнаружил осцилляции этих сигналов. Они напоминали электрическую активность сокращающегося сердца и, по его мнению, свидетельствовали о пульсациях паренхимных клеток. Осцилляции практически отсутствовали в эпидермисе стебля, но быстро нарастали по мере проникновения зонда внутрь стебля и оказывались максимальными у самого внутреннего слоя клеток, граничившего непосредственно с сосудистой тканью (рис. 14).
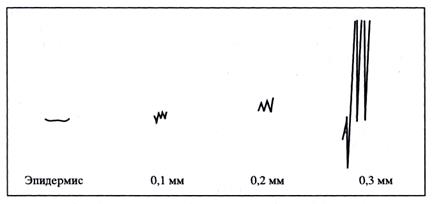 |
Рис. 14. Запись электрических сигналов клеток стебля по мере углубления в него регистрирующего зонда: на расстоянии 0,3 мм от поверхности стебля амплитуда сигналов резко увеличивается. Это соответствует внутреннему слою клеток коры, граничащего с молодой сосудистой тканью.
Под действием паров хлороформа осцилляции постепенно исчезали, то есть они зависели от дыхания. Прямой контакт зонда с ксилемой не давал уже никакого сигнала. По мнению Боса, это указывало на то, что сами сосуды ксилемы не принимают активного участия в поднятии воды. вода поднимается вверх благодаря перистальтическим сокращениям паренхимных клеток, окружающих сосуды.
«У двудольных растений, писал Д.Ч. Бос, 1964, пульсирующая ткань представляет собой цилиндрическую трубку, которая тянется по всей длине стебля. Функция цилиндра … похожа на функцию продолговатого сердца низших животных, осуществляющих циркуляцию крови волнами перистальтических движений. Продвижение соков в растениях – простейший процесс перистальтики, где волна сокращения выталкивает сок вперед. Ни листья, ни корни не являются обязательно необходимыми для восхождения соков … движение соков продолжается и после полного удаления корней и листьев».
В пользу справедливости представлений Д.Ч. Боса свидетельствуют опыты с Heracleum, у которого сосуды в ксилеме окружает множество паренхимных клеток. Поэтому ксилема дышит ни чуть не слабее флоэмы. Следовательно, энергии, необходимой для сокращений клеток, окружающих ксилемные сосуды, вполне достаточно.
Несмотря на то, что Бос был сторонником представления об активном участии паренхимных клеток в транспортных процессах, он не отрицал и роль присасывающего действия транспирации в поднятии воды. То есть он был сторонником двухкомпонентной движущей силы транспорта воды в стебле. По его мнению «во время интенсивной транспирации физиологическая пульсация не достаточна по сравнению с потребностью… Следовательно, действуют два фактора – физиологическая пульсация, производимая активными клетками вдоль всей коры, и физическая передача воды вдоль … ксилемы».
Часть восходящего тока направляется, помимо сосудистой системы стебля непосредственно по живым клеткам, через межклеточные контакты (то есть по симпласту). Доля этого потока составляет 1 – 10% общего потока.
Флоэмный транспорт в стебле также связан с микроперистальтическими движениями элементов флоэмы и обнаруживает импульсную ритмичность (Пинхасов, 1981; Фенсом и др., 1994). Причем Флоэмный и ксилемный потоки, по-видимому, асинхронны.








