2015-07-03
2015-07-03 6662
6662У більшості рослин (дводольних і голонасінних) первинна будова кореня зберігається недовго і переходить у вторинну будову. Така перебудова пов'язана з утворенням на певному етапі їхнього розвитку (після появи перших листків) у центральному циліндрі кореня вторинної меристеми — камбію. За рахунок клітин камбію утворюються вторинні елементи ксилеми і флоеми. У дерев і кущів вторинна ксилема і флоема наростають кільцями, тому будова кореня подібна до будови стебла. Первинна кора та ендодерма поступово відмирають і злущуються, а з перициклу утворюється перидерма. Нові шари перидерми закладаються у глибших шарах вторинної флоеми. Так поступово виникає вторинна будова кореня.
Вторинна будова кореня хвойних ы дводольних рослин характерна для тих ділянок кореня, які виконують функцію зміцнення і закріплення рослини в грунті, а не всмоктування поживних речовин (мал. 2). Із зміною функції змінюється й анатомічна будова кореня. Вторинні зміни відбуваються з появою камбію, що закладається в паренхімі центрального циліндра, яка розмежовує первинні флоему і ксилему. Клітини паренхіми діляться тангентально і утворюють камбіальні дуги, які своїми кінцями упираються в перицикл. Ділянки перициклу, які замкнені між кінцями камбіальних дуг, добудовують кільце камбію, яке має неправильну форму. Отже, цей камбій має паренхімно-перициклічне походження. Камбій, що виникає за рахунок паренхіми центрального циліндра, до центру відкладає вторинну ксилему, яка в сформованому стані складається з великих судин, мало потовщених трахеїд і лібриформу, потужно розвиненої тонкостінної ксилемної паренхіми. Відкладаючись у великій кількості, ксилема відтісняє камбій і він набуває правильної кільцевої форми. До периферії камбій відкладає вторинну флоему, яка складається із ситоподібних трубок, клітин-супутниць, камбієформу, флоемної паренхіми. Камбій перициклічного походження відкладає паренхіму радіальних променів, які розділяють ділянки вторинних ксилеми і флоеми. Внаслідок наростання вторинних ксилеми і флоеми первинна ксилема відтісняється до центру, а первинна флоема відтісняється вторинною флоемою до периферії. В рослин пучкового типу будови виникають відкриті колатеральні провідні пучки.
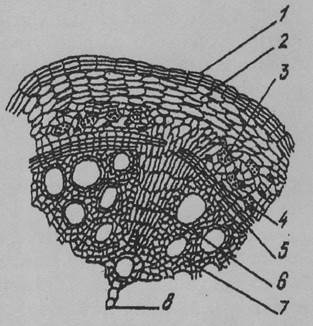
Рис. 2. Вторинна будова кореня: 1 — перидерма, 2 — паренхіма кори, 3,7 — вторинна ксилема, 4 — міжпучковий камбій, 5 — пучковий камбій, 6 — радіальний промінь, 8 — первинна ксилема
Зміни в корені торкаються не лише центрального циліндра, а і його периферії. Клітини перициклу, що не брали участі в утворенні пучкового камбію, діляться і перетворюються в клітини фелогену. Фелоген £ свою чергу утворює до периферії клітини корку, а всередину — клітини фелодерми. Корок, фелоген і фелодерма разом узяті називаються перидермою. Перидерма ізолює первинну кору від центрального циліндра, відбувається процес злущування, або "линяння" кореня, який підсилюється значним потовщенням центрального циліндра за рахунок розростання вторинних елементів флоеми і ксилеми. Після злущування первинної кори на її місці виникає вторинна кора і формується вторинна будова з дуже розвиненими вторинними ксилемою і флоемою, паренхімою кори, радіальними променями, пучковим і міжпучковим камбієм, скорковілими клітинами перидерми. Від первинної будови залишається тільки первинна ксилема в центрі і первинна флоема, відтіснена до периферії вторинною флоемою.
Корені моркви і редьки мають типову вторинну будову (рис.), але в зв'язку із запасанням поживних речовин вони дуже потовщуються. В моркви корінь потовщується за рахунок надмірного розростання паренхіми, в якій розкидані в радіальному напрямі ділянки флоеми. Вторинна ксилема слабо розвинена.
Коренеплід редьки характеризується надмірним розростанням вторинної ксилеми, особливо ксилемної паренхіми. За ксилемою розміщується камбій, який до периферії відчленовує вторинну флоему. Вторинна флоема слабо розвинена. За нею знаходяться ділянка первинної флоеми, паренхіма кори і перидерма.
Третинна будова коренявластива буряку. В процесі онтогенезу у нього спочатку розвивається первинна будова кореня. В центрі її міститься двопроменева ксилема, а по боках первинна флоема, відмежована прошарком паренхіми центрального циліндра. Перицикл одношаровий. Далі розміщена первинна кора з великими паренхімними клітинами.
У папоротей і однодольних первинна будова зберігається в коренів упродовж усього життя. У голонасінних і дводольних вона змінюється вторинною будовою, яку можна спостерігати в провідній зоні.
Спочатку в коренях дводольних камбію немає, він виникає перед вторинними змінами і формує їх. Камбій закладається дугами під променями первинної флоеми, утворюється з розташованої там паренхіми. При цьому її клітини витягуються і поділяються тангентальними перегородками. Оскільки центр кореня займає первинна ксилема, камбіальна дуга опиняється між ксилемою і флоемою. До першої вона повернена своїм опуклим боком, до другої — угнутим. Далі камбіальні дуги подовжуються уздовж променів ксилеми, доходячи до перициклу. Клітини перициклу також починають ділитися, утворюючи камбій. Камбіальне кільце замикається, однак форма його виявляється лопатевою (мал. 3). •
Камбіальні клітини починають працювати в дугах під променями флоеми ще до замикання повного кільця. Звичайно, вторинної деревини утворюється більше, ніж лубу, і камбіальне кільце поступово випрямляється. Зрозуміло, що воно неоднорідне за походженням: одні його ділянки виникли з паренхіми провідного пучка, інші — з перициклу. Функціонують ці ділянки також по-різному: похідні паренхіми утворюють елементи вторинного лубу і вторинної деревини, а похідні перициклу — паренхімну тканину. Іншими словами, меристема, утворена з паренхіми, працює як пучковий камбій, а утворена з перициклу, — як міжпучковий.
У центрі кореня залишається первинна ксилема. Тяжі паренхіми йдуть від променів первинної ксилеми, розсікаючи вторинну деревину і вторинний луб. Вони аналогічні серцевинним променям стебла, але, оскільки в коренях немає серцевини, тут їх називають паренхімними променями. Паренхімні промені, що тягнуться від первинної ксилеми, називають первинними. Але утворюються і вторинні промені. Вони проходять у вторинній деревині й лубі та коротше і вужче за первинні. У зв'язку з тим, що первинні паренхімні промені досить широкі, що формується у корені при вторинних змінах утворюється пучкова будова. Причому пучки, що виникли, є колатеральними і відкритими (мал. 3).
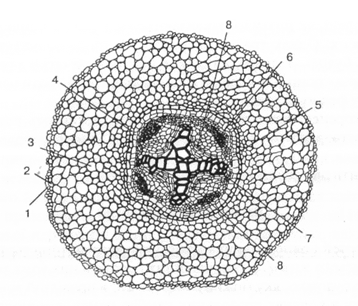
Мал. 3. Закладення камбію в корені дводольних (кінські боби):
1 — епіблема; 2 — екзодерма; З — мезодерма; 4 — ендодерма;
5 — перицикл; 6— флоема; 7— ксилема; 8— камбій
Первинна флоема при формуванні вторинної структури кореня відтискується вторинним лубом далеко на периферію, де втрачає свою функцію і сплющується. У коренях унаслідок сезонних змін утворюються річні кільця, але вони більш вузькі, ніж у стеблах, і межі між ними виражені слабко.
У формуванні вторинної будови кореня бере участь не тільки камбій, що утворює луб і деревину, але і корковий камбій.
Ще в процесі формування камбіальних дуг перицикл вичленяє назовні, у бік первинної кори, кільце фелогену, що утворює перидерму. Первинна кора, ізольована шаром непроникного корка від провідних тканин, відмирає і під тиском деревини, що розростається, тріскається і скидається. Тому первинна кора у вторинній будові кореня відсутня. Замість неї під перидермою може знаходитися луб'яна паренхіма, а також паренхіма, яку утворює фелоген до формування ним перидерми. У старих коренів перидерма може замінюватися кіркою.
Вторинна будова кореня відрізняється від такої стебла цілим рядом особливостей.
1. У корені більш широкопросвітні й тонкостінні елементи вторинної ксилеми і розташовані вони щільніше.
2. Клітини паренхіми кореня більші за розміром, а складені з них паренхімні промені значно ширші за серцевинні промені стебла.
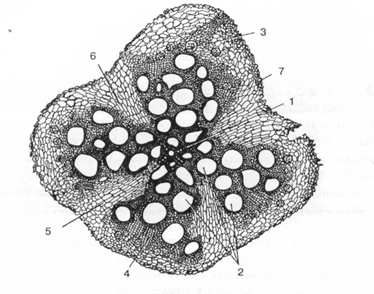
Рис. 4. Вторинна будова кореня гарбуза:
1 — первинна ксилема; 2 — вторинна ксилема; 3 — вторинна флоема; 4 — пучковий камбій; 5 — міжпучковий камбій;
6 — паренхімні промені; 7 — перидерма
3. У коренях мало механічної тканини.
4. У них відсутня первинна кора.
5. Річні кільця тонші і виявляються нечітко.
6. Судини в коренях рідко закупорюються тилами.
Розходження в анатомічній будові основних осьових органів рослин — стебла і кореня пояснюються тим, що вони виконують різні функції, а також існуванням їх у різних середовищах — повітряному і ґрунтовому [4; c. 15].