 2015-08-13
2015-08-13 519
519| Активность, % |
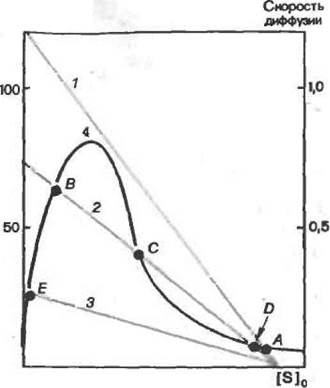
| лнзованного фермента, подверженного ни- гнбнрованию субстратом: прямые tf 2 и.? —зависимости скорости диффузии суб- стРата к ФеРмен^ «[51* ч>™и-.зависимость скорости ферментативной ре- акции от концентрации субстрата в от- сутствие диффузионных ограничений; точ кн И. й. t", £>, f)—стационарные со- стояния- Kor«a скорость диффузии суб- страта к фераденту равна скорости для иммобилизованного фермента |
 субстрата определяется двумя взаимосвязанными факторами: степенью диффузионных ограничений субстрата и его концентрацией в свободном растворе. При этом- чем выше концентрация субстрата н чъм ниже исходная активность фермента, тем больше вероятность, что ферментативная реакция будет протекать в кинетическом режиме. Поэтому понятно, что увеличение концентрации субстрата, сопровождаемое снижением ферментативной активности {за счет ннгибнровання фермента), может вызвать резкий переход реакции им диффузионного режима в кинетический.
субстрата определяется двумя взаимосвязанными факторами: степенью диффузионных ограничений субстрата и его концентрацией в свободном растворе. При этом- чем выше концентрация субстрата н чъм ниже исходная активность фермента, тем больше вероятность, что ферментативная реакция будет протекать в кинетическом режиме. Поэтому понятно, что увеличение концентрации субстрата, сопровождаемое снижением ферментативной активности {за счет ннгибнровання фермента), может вызвать резкий переход реакции им диффузионного режима в кинетический.
Рассмотрим этот вывод подробнее. Согласно первому закону Фика скорость переноса субстрата к ферменту прямо пропорциональна концентрации субстрата у поверхности носителя. Катализируемая иммобилизованным ферментом реакция ' достигает стационарного состояния, когда скорость переноса субстрата к ферменту становится равной скорости его ферментативной трансформации. На рис. 22 стационарные состояния задаются точками пересечения кривой зависимости ферментативной активности от концентрации субстрата с прямой скорости диффузии субстрата к ферменту. При высоких скоростях диффузии, т. е. незначительных диффузионных ограничениях (прямая 1), так и при низких скоростях диффузии, т. е. весьма больших диффузионных ограничениях {прямая 3), ня рис. 22 наблюдается лишь одна точка пересечения — одно стационарное состояние. При промежуточных скоростях диффузии (прямая 2) имеется три точки Пересе-чения, т.е. допустимы три различных стационарных состояния. Если построить график зависимости реальной скорости ферментативной реакции от концентрации субстрата, то при высоких и низких значениях концентрации субстрата возможно только одно стационарное состояние, а при промежуточных значениях — три состояния: А, В и С. Таким образом, получаем гистерезисную
]\5

|
| Рис. 23. Гнстереэисная зависимость реальной скорости реакции для иммобилизован ного фермента, подверженного ингибирсша-нию субстратом, при диффузионных ограничениях субстрата от его концентрации; стрелки обозначено направление изменения концентрации субстрата [SJo в системе |
кривую (рис. 23), Она означает, что скорость ферментативной реакции определяется не только концентрацией субстрата во внешнем растворе, но и зависит от направления изменения концентрации субстрата. Так, при повышении концентрации субстрата во внешнем растворе скорость ферментативной реакции увеличивается. Однако при достижении некоторой критической концентрации субстрата система переходит из диффузионной области в кинетическую и скорость реакции суще-ственно падает {рис. 23). При понижении концентрации субстрата скорость реакции остается низкой до тех пор пока не произойдет обратный переход из кинетической области в диффузионную. При дальнейшем уменьшении концентрации субстрата скорость ферментативной реакции снова начнет падать.
Причины того же характера приводят к появлению внутри-мембранных колебательных процессов. В качестве одного из примеров приведем катализируемый папаином, иммобилизованным в искусственной мембране, гидролиз этилового эфира 1^-бензойл-и-аргинина. Один нз продуктов ферментативной реакции — аминокислота, накапливается в мембране (за счет ограничения диффузии) и тем самым с-двигает рН внутри мембраны п сторону более кислых значений. Это неизбежно уменьшает гидролитическую активность лапаина, так как фермент «выходит» из своего рН-оптимума активности. В дальнейшем за счет более быстрой диффузии Н+ по сравнению с ОЬГвозрастает значение рН внутри мембраны и активность лапаина снова увеличивается. Таким образом, был получен микрореактор с периодом колебания 20 с. Изменением концентрации фермента в мембране, толщины мембраны и других параметров системы удается варьировать продолжительность одного колебания.
Анализ условий, при которых могут наблюдаться гистерезис-иые и колебательные явления, показал, что для их возникновения необходимо» чтобы, во-первых, на зависимости скорости ферментативной peaк г inи от концентрации субстрата имелся максимум или точка перегиба, т, е, фермент активируется или ингибируется








