2015-10-16
2015-10-16 1188
1188Спинной мозг находится в позвоночном канале, его масса от 34 до 38 г. Он представляет собой тяж, слегка сплюснутый в переднезаднем направлении, протяженностью 41-45 см. Соотношение между длиной спинного мозга и позвоночного канала меняется в процессе развития организма. У трехмесячного плода длина спинного мозга равна длине позвоночного канала, однако в последующем удлинение спинного мозга отстает от увеличения длины позвоночника и его канала. У новорожденного спинной мозг заканчивается на уровне III поясничного позвонка, у взрослого человека нижний конец (конус) спинного мозга (conus medullaris) находится на уровне верхнего края II поясничного позвонка; здесь спинной мозг переходит в конечную нить {ftlum terminate), спускающуюся в крестцовый отдел позвоночного канала и фиксирующуюся здесь к надкостнице обычно II крестцового позвонка (рис. 2.8).
Спинной мозг имеет 31—32 пары передних и столько же пар задних корешков. Участок спинного мозга, точнее его серого вещества, анатомически и функционально связанный с парой передних и парой задних корешков, называется сегментом. 8 верхних сегментов образуют шейный отдел спинного мозга, грудной отдел его состоит из 12 сегментов, поясничный и крестцовый отделы имеют по 5 сегментов каждый, последние 1-2 сегмента составляет копчиковый отдел.
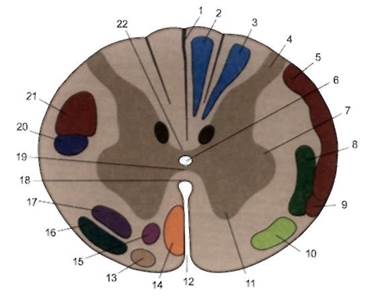
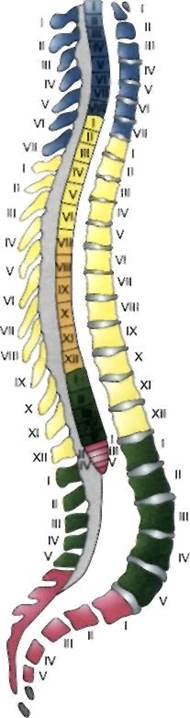 | Рис. 2.8. Топографические соотношения между сегментами спинного мозга и позвонками. Шейные сегменты спинного мозга принято обозначать латинской буквой С (cervicalis) с указанием порядкового номера сегмента (например, V шейный сегмент — Cv). Грудные сегменты обозначаются буквами Th (thoracalis), Поясничные — L (lumbalis) Крестцовые — Со (coccygea) также в сочетании с порядковым номером соответствующего сегмента. Каждый сегмент спинного мозга через посредство своих корешков обеспечивает иннервацию определенного участка тела, именуемого метамером, при этом входящие в метамер мышцы составляют миотом, кости — склеротом, а относящийся к одному метамеру участок кожи называется дерматомом. Название каждого миотома, склеротома и дерматома определяется в соответствии с названием соответствующего ему сегмента спинного мозга Толщина спинного мозга не идентична на уровне разных его отделов. На среднем грудном уровне — приблизительно 10 мм. На уровне сегментов, обеспечивающих иннервацию конечностей, спинной мозг утолщен до 13-16 мм в диаметре. Эти утолщения находятся на уровнях Cv—Thn (шейное утолщение — intumescentia cervicalis) и Ln~Sn (пояснично-крестцовое утолщение — intumescentia lumbosacral). На поперечном срезе спинного мозга видно, что его центральную часть занимает серое вещество, образующее фигуру, несколько напоминающую бабочку. Ее форма варьирует на разных уровнях спинного мозга. В ней выделяются центральная часть серого вещества, два задних и два передних рога, а на уровне CVI|i—LUI — еще и боковые рога спинного мозга. Белое вещество на поперечном срезе спинного мозга занимает не заполненные серым веществом участки. Оно образует передние, задние и латеральные канатики (столбы) спинного мозга, состоящие из нервных волокон, формирующих восходящие (афферентные) и нисходящие (эфферентные) проводящие пути (см. главу 8). |
СПИННОЙ МОЗГ И СПИННОМОЗГОВЫЕ НЕРВЫ 8.1, ОБЩИЕ ПОЛОЖЕНИЯ В предыдущих главах (см. главы 2, 3, 4) были рассмотрены общие принципы строения спинного мозга и спинномозговых нервов, а также проявления чувствительной и двигательной патологии при их поражении. В этой главе уделено внимание главным образом частным вопросам морфологии, функции и некоторым формам поражения спинного мозга и спинномозговых нервов. 8.2. СПИННОЙ МОЗГ Спинной мозг — часть центральной нервной системы, сохранившая отчетливые черты сегментарного строения, свойственные прежде всего его серому веществу. Спинной мозг имеет многочисленные взаимные связи с головным мозгом. Оба этих отдела ЦНС в норме функционируют как единое целое. У млекопитающих, в частности у человека, сегментарная деятельность спинного мозга постоянно испытывает на себе влияние эфферентных нервных импульсов, исходящих из различных структур головного мозга. Это влияние в зависимости от многих обстоятельств может быть активизирующим, облегчающим или тормозящим. 8.2.1. Серое вещество спинного мозга Серое вещество спинного мозга составляют главным образом тела нервных и глиальных клеток. Неидентичность количества их на разных уровнях спинного мозга обусловливает вариабельность объема и конфигурации серого вещества. В шейном отделе спинного мозга передние рога широкие, в грудном отделе серое вещество на поперечном срезе становится похожим на букву «Н», в по-яснично-крестцовом отделе особенно значительны размеры как передних, так и задних рогов. Серое вещество спинного мозга фрагментируется на сегменты. Сегментом является фрагмент спинного мозга, анатомически и функционально связанный с одной парой спинномозговых нервов. Передние, задние и боковые рога можно рассматривать как фрагменты вертикально расположенных столбов — переднего, заднего и бокового, отделенных друг от друга состоящими из белого вещества канатиками спинного мозга. В осуществлении рефлекторной деятельности спинного мозга важную роль играет следующее обстоятельство: практически все аксоны клеток спинномозговых узлов, входящие в спинной мозг в составе задних корешков, имеют ответвления — коллатерали. Коллатерали чувствительных волокон контактируют непосредственно с периферическими мотонейронами, расположенными в передних рогах, или с вставочными нейронами, аксоны которых также достигают тех же двигательных клеток. Коллатерали аксонов, отходящих от клеток межпозвонковых узлов, не только доходят до соответствующих периферических мотонейронов, расположенных в передних рогах ближайших сегментов спинного мозга, но и проникают в соседние его сегменты, формируя при этом так называемые спинно-спинальные межсегментарные связи,обеспечивающие иррадиацию возбуждения, пришедшего в спинной мозг после раздражения расположенных на периферии рецепторов глубокой и поверхностной чувствительности. Этим объясняется распространенная рефлекторная двигательная реакция в ответ на локальное раздражение.Такого рода явления особенно типичны при уменьшении тормозящего влияния пирамидных и экстрапирамидных структур на периферические мотонейроны, входящие в состав сегментарного аппарата спинного мозга. Нервные клетки,составляющие серое вещество спинного мозга, по своей функции могут быть разделены на следующие группы: 1. Чувствительные клетки (Т-клетки задних рогов спинного мозга) являются телами вторых нейронов чувствительных путей. Большая часть аксонов вторых нейронов чувствительных путей в составе белой спайки переходит напротивоположную сторону, где участвует в формировании боковых канатиков спинного мозга, образуя в них восходящие спиноталамические пути и передний спиномозжечковый тракт Говерса. Аксоны вторых нейронов, не перешедшие на противоположную сторону, направляются в гомолатеральный боковой канатик и формируют внем задний спиномозжечковый путь Флексига. 2. Ассоциативные (вставочные) клетки, относящиеся к собственному аппарату спинного мозга, участвуют в формировании его сегментов. Их аксоны заканчиваются в сером веществе тех же или близко расположенных спиналь-ных сегментов. 3. Вегетативные клетки расположены в боковых рогах спинного мозга на уровне С8— L2 сегментов (симпатические клетки) ив сегментах S3--S5 (парасимпатические клетки). Аксоны их покидают спинной мозг в составе передних корешков. 4. Двигательные клетки (периферические мотонейроны) составляют передние рога спинного мозга. К ним сходится большое количество нервных импульсов, идущих из различных отделов головного мозга по многочисленным нисходящим пирамидным и экстрапирамидным путям. Кроме того, нервные импульсы к ним приходят по коллатералям аксонов псевдоуниполярных клеток, тела которых находятся в спинномозговых узлах, а также по коллатералям аксонов чувствительных клеток задних рогов и ассоциативных нейронов того же или других сегментов спинного мозга, несущих информацию главным образом от рецепторов глубокой чувствительности, и по аксонам, расположенным в передних рогах спинного мозга, клеток Реншоу, которые посылают импульсы, снижающие уровень возбуждения альфа-мотонейронов и, следовательно, уменьшающие напряжение поперечнополосатых мышц. Клетки передних рогов спинного мозга служат местом интеграции возбуждающих и тормозных импульсов, поступающих от различных источников. Сложение приходящих в мотонейрон возбуждающих и тормозных биопотенциалов определяет его суммарный биоэлектрический заряд и в связи с этим особенности функционального состояния. Среди периферических мотонейронов, расположенных в передних рогах спинного мозга, выделяются клетки двух видов: а) альфа-мотонейроны — крупные двигательные клетки, аксоны которых имеют толстую миелиновую оболочку (волокна А-альфа) и заканчиваются в мышце концевыми пластинками; они обеспечивают степень напряжения экстрафузальных мышечных волокон, составляющих основную массу поперечнополосатых мышц; б) гамма-мотонейроны — мелкие двигательные клетки, аксоны которых имеют тонкую миелиновую оболочку (волокна А-гамма) и, следовательно, меньшую скорость проведения нервных импульсов. Гамма-мотонейроны составляют приблизительно 30% от всех клеток передних рогов спинного мозга; аксоны их направляются к интрафузальным мышечным волокнам, входящим в состав проприо-рецепторов — мышечных веретен. Мышечное веретено состоит из нескольких тонких интрафузальных мышечных волокон, заключенных в веретенообразную соединительнотканную капсулу. На интрафузальных волокнах заканчиваются аксоны гамма-мотонейронов, влияющих на степень их напряжения. Растяжение или сокращение интрафузальных волокон ведет к изменению формы мышечного веретена и к раздражению спиралевидного волокна, окружающего экватор веретена. В этом волокне, являющемся началом дендрита псевдоуниполярной клетки, возникает нервный импульс, который направляется к телу этой клетки, находящемуся в спинномозговом ганглии, а затем по аксону той же клетки — в соответствующий сегмент спинного мозга. Конечные ветви этого аксона непосредственно или через вставочные нейроны достигают альфа-мотонейрона, оказывая на него возбуждающее или тормозное влияние. Таким образом, с участием гамма-клеток и их волокон создается гамма-петля, обеспечивающая поддержание тонуса мышцы и фиксированное положение определенной части тела или сокращение соответствующих мышц. Кроме того, гамма-петля обеспечивает трансформацию рефлекторной дуги в рефлекторное кольцо и принимает участие в формировании, в частности, сухожильных, или миотатических, рефлексов. Моторные нейроны в передних рогах спинного мозга формируют группы, каждая из которых иннервирует мышцы, объединенные общностью функций. По длиннику спинного мозга располагаются передневнутренние группы клеток передних рогов, обеспечивающие функцию мышц, влияющих на положение позвоночного столба, и передненаружные группы периферических мотонейронов, от которых зависит функция остальных мышц шеи и туловища. В сегментах спинного мозга, обеспечивающих иннервацию конечностей, имеются дополнительные группы клеток, располагающиеся преимущественно позади и снаружи уже упомянутых клеточных объединений. Эти дополнительные группы клеток являются основной причиной шейного (на уровне C5—Th2 сегментов) и поясничного (на уровне L2—S2 сегментов) утолщений спинного мозга. Они обеспечивают главным образом иннервацию мышц верхних и нижних конечностей. Двигательная единица нейромоторного аппарата состоит из нейрона, его аксона и иннервируемой им группы мышечных волокон. Сумма периферических мотонейронов, принимающих участие в иннервации одной мышцы, известна как ее двигательный пул, при этом тела мотонейронов одного двигательного пула могут располагаться в нескольких соседних сегментах спинного мозга. Возможность поражения части двигательных единиц, входящих в состав мышечного пула, является причиной частичного поражения иннервируемой им мышцы, как это бывает, например, при эпидемическом полиомиелите. Распространенное поражение периферических мотонейронов характерно для спинальных амиотрофий, относящихся к наследственным формам нервно-мышечной патологии. Среди других заболеваний, при которых в спинном мозге избирательно поражается серое вещество, следует отметить сирингомиелию. Сирингомиелия характеризуется расширением обычно редуцированного центрального канала спинного мозга и формированием глиоза в его сегментах, при этом чаще страдают задние рога, и тогда в соответствующих дерматомах возникает расстройство чувствительности по диссоциированному типу. Если дегенеративные изменения распространяются также на передние и боковые рога, в метамерах тела, одноименных пораженным сегментам спинного мозга, возможны проявления периферического пареза мышц и вегетативно-трофические нарушения. В случаях гематомиелии (кровоизлиянии в спинной мозг), обычно возникающей вследствие травмы спинного мозга, симптоматика имеет сходство с сирингомиелитическим синдромом. Поражение при травматическом кровоизлиянии в спинной мозг преимущественно серого вещества объясняется особенностями его кровоснабжения. Серое вещество является также местом преимущественного формирования интрамедуллярных опухолей, растущих из его глиальных элементов. В дебюте опухоли могут проявиться симптомами поражения определенных сегментов спинного мозга, но в последующем в процесс вовлекаются медиальные отделы прилежащих канатиков спинного мозга. В этой стадии роста интрамедулляр-ной опухоли несколько ниже уровня ее локализации появляются нарушения чувствительности по проводниковому типу, которые в последующем постепенно спускаются вниз. Со временем на уровне расположения интрамедуллярной опухоли может развиться клиническая картина поражения всего поперечника спинного мозга. Признаки сочетанного поражения периферических мотонейронов и корти-ко-спинальных проводящих путей характерны для бокового амиотрофическо-го склероза (синдром БАС). В клинической картине при этом возникают различные комбинации проявлений периферического и центрального пареза или паралича. В таких случаях по мере гибели все большего числа периферических мотонейронов симптомы уже развившегося центрального паралича сменяются проявлениями паралича периферического, которые со временем все более преобладают в клинической картине заболевания. 8.2.2. Белое вещество спинного мозга Белое вещество формирует расположенные по периферии спинного мозга канатики, состоящие из восходящих и нисходящих проводящих путей, большинству из которых уже уделялось внимание в предыдущих главах (см. главы 3, 4). Теперь можно дополнить и обобщить изложенные там сведения. Нервные волокна, имеющиеся в спинном мозге, можно дифференцировать на эндогенные, являющиеся отростками собственных клеток спинного мозга, и экзогенные — состоящие из проникших в спинной мозг отростков нервных клеток, тела которых расположены в спинномозговых узлах или входят в состав структур головного мозга. Эндогенные волокна могут быть короткими и длинными. Чем короче волокна, тем ближе к серому веществу спинного мозга они расположены. Короткие эндогенные волокна формируют спиноспинальные связи между сегментами самого спинного мозга (собственные пучки спинного мозга — fasciculi proprii). Из длинных эндогенных волокон, являющихся аксонами вторых чувствительных нейронов, тела которых расположены в задних рогах сегментов спинного мозга, сформированы афферентные пути, проводящие импульсы болевой и температурной чувствительности, идущие к таламусу, и импульсы, направляющиеся к мозжечку (спиноталамические и спиномозжечковые пути). Экзогенные волокна спинного мозга являются аксонами клеток, находящихся за его пределами. Они могут быть афферентными и эфферентными. Афферентные экзогенные волокна составляют тонкий и клиновидный пучки, формирующие задние канатики. Среди эфферентных проводящих путей, состоящих из экзогенных волокон, следует отметить боковой и передний корти-ко-спинальные тракты. Из экзогенных волокон состоят также относящиеся к экстрапирамидной системе красноядерно-спинномозговой, преддверно-спин-номозговой, оливо-спинномозговой, покрышечно-спинномозговой, вестибу-ло-спинномозговой, ретикуло-спинномозговой проводящие пути. В канатиках спинного мозга наиболее важные проводящие пути распределены следующим образом (рис. 8.1): Задние канатики (funiculus posterior seu dorsalis) состоят из восходящих путей, проводящих импульсы проприоцептивной чувствительности. В нижней части спинного мозга задний канатик составляет тонкий пучок Голая (fasciculus gracilis). Начиная от среднегрудного отдела спинного мозга и выше, латераль-нее тонкого пучка, формируется клиновидный пучок Бурдаха (fasciculus cuneatus). В шейном отделе спинного мозга оба этих пучка хорошо выражены и разделены глиальной перегородкой. Поражение задних канатиков спинного мозга ведет к нарушению проприоцептивной и к возможному снижению тактильной чувствительности ниже уровня поражения спинного мозга. Проявлением такой формы патологии является нарушение в соответствующей части тела обратной афферентации в связи с отсутствием должной информации, направляющейся в головной мозг, о положении частей тела в пространстве. В результате возникают сенситивная атаксия и афферентный парез, при этом характерны также гипотония мышц и сухожильная гипорефлексия или арефлексия. Такая форма патологии характерна для спинной сухотки, фуникулярного миелоза, входит в состав симпто-мокомплексов, характерных для различных форм спиноцеребеллярной атаксии, в частности атаксии Фридрейха. Боковые канатики (funiculus lateralis) состоят из восходящих и нисходящих путей. Дорсолатеральный отдел бокового канатика занимает задний спино-мозжечковый путь Флексига (tractus spinocerebellaris dorsalis). В вентролате-ральном отделе находится передний спиномозжечковый путь Говерса (tractus spinocerebellaris ventralis). Медиальнее пути Говерса располагается путь импульсов поверхностной чувствительности — латеральный спиноталамический путь (tractus spinothalamicus lateralis), позади него красноядерно-спинномозговой путь (tractus rubrospinalis), между ним и задним рогом — латеральный корко-во-спинномозговой (пирамидный) путь (tractus corticospinalis lateralis). Кроме того, в боковом канатике проходят спинно-ретикулярный путь, покрышечно-спинномозговой путь, оливоспинномозговой путь, а вблизи серого вещества рассеяны вегетативные волокна.
Так как в боковом канатике корково-спинномозговой путь находится дор-сальнее латерального спиноталамического пути, то поражение заднего сегмента спинного мозга может привести к расстройству глубокой чувствительности в сочетании с пирамидным нарушением ниже уровня локализации патологического очага при сохранности поверхностной чувствительности (синдром Русси—Лермитта— Шельвена). Избирательное поражение входящих в состав боковых канатиков спинного мозга пирамидных путей возможно, в частности, при семейной спастической параплегии, или болезни Штрюмпеля, при которой, кстати, в связи с неоднородностью составляющих пирамидный путь волокон характерно расщепление пирамидного синдрома, что проявляется нижним спастическим парапарезом с преобладанием спастического напряжения мышц над снижением их силы. Передние канатики (funiculus anterior seu ventralis) состоят в основном из эфферентных волокон. К срединной щели прилежит покрышечно-спинномозговой проводящий путь (tractus tectospinalis), относящийся к системе нисходящих экстрапирамидных путей. Латеральнее расположены передний (неперекре-щенный) корково-спинномозговой (пирамидный) путь (tractus corticospinalis anterior), преддверно-спинномозговой путь (tractus vestibulospinalis), передний ретикулярно-спинномозговой путь (tractus reticulospinalis anterior) и афферентный передний спиноталамический путь (tractus spinothalamicus anterior). Позади них проходит медиальный продольный пучок (fasciculis longitudinalis medialis), несущий импульсы от ряда клеточных образований покрышки ствола. При развитии ишемии в бассейне передней спинномозговой артерии (синдром Преображенского) нарушается кровообращение в передних 2/3 поперечника спинного мозга. На уровне зоны ишемии развивается вялый паралич мышц, ниже этого уровня — спастический. Характерны также расстройство болевой и температурной чувствительности по проводниковому типу и нарушение функции тазовых органов. Проприоцептивная и тактильная чувствительность при этом сохраняется. Этот синдром описал в 1904 г. МА. Преображенский (1864-1913). |