 2015-10-16
2015-10-16 535
535АмплшпуЗа ПОК (первой негативной волны)
Интенсивность ощущения
< -------------0,5 сек--------------->■-----------------------------—
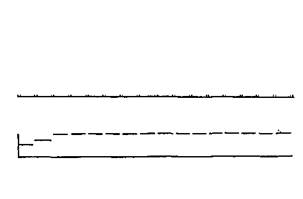
Рис. VI-6. Схема взаимоотношений между последовательностью прямоугольных импульсов (пороговой интенсивности и длительностью 0,5 мсек), приложенных к постцентральной извилине, и амплитудой прямых ответов коры (ПОК), записанных вблизи области раздражения. Третья линия показывает, что субъективные ощущения не возникают примерно в течение первых 0,5 сек стимуляции. Интенсивность появившегося после этого периода ощущения сохраняется на постоянном уровне, несмотря на то, что стимуляция продолжается (Libet, 1966).
положения тела в пространстве могут быть вызваны и при отсутствии каких-либо заметных изменений в самом теле, которые обычно переживаются пациентом как ощущения. Либет показал, что ощущение, вызываемое стимуляцией, возникает не сразу: проходит минимум полсекунды и максимум 5 мин, прежде чем пациент начинает что-либо ощущать. Видимо, электрическая стимуляция должна создать некоторое состояние в мозговой ткани и, только когда это состояние возникает, пациент начинает что-то чувствовать (рис. VI-6).
ВОСПРИЯТИЕ
Сформулированная в этой главе гипотеза имеет во многих отношениях решающее значение для последовательно развиваемой здесь точки зрения на функции нервной системы. Но она не бесспорна. В первой части книги был дан обзор логических операций, которые выполняются нервной системой. Здесь я хочу показать, каким образом эти операции делают возможным осознание организмом того, что происходит в данный момент и в данном месте, иначе говоря, как формируется восприятие настоящего.
Проблема состоит в следующем: если вы посмотрите на своего друга, а затем на его соседа, то вы сразу же понимаете, чем они отличаются друг от друга. Далее происходит небольшая интерфе-
ренция — если только она действительно происходит — между тем, что вы видели в первый момент, и тем, что было воспринято в следующий. При слуховой форме коммуникации такое непрерывное быстрое узнавание — музыкальных фраз, фонетических сочетаний речи и т. д. — обычное явление. Традиционный взгляд на функции нервной системы сталкивается со значительными трудностями при объяснении непосредственности, точности и видимой многомерности мимолетных восприятий. Здесь, должно быть, действует единый процесс. Но каким образом он действует?
Д. Хебб детально рассмотрел эту проблему в первых трех главах своего классического труда «Организация поведения» (1949). Он пишет: «Следует решить, зависит ли восприятие 1) от возбуждения специфических клеток или 2) от структуры возбуждений, местоположение которой не имеет значения». Хебб сделал следующий вывод: «Восприятие зависит от возбуждения специфических клеток в какой-то точке центральной нервной системы».
Когда накопились нейрофизиологические данные, особенно полученные в микрозлектродных исследованиях Юнга (1961), Маунткасла (1957); Матураны, Леттвипа, Мак-Каллока и Питтса (1960); Хьюбела и Визела (1962), точка зрения Хебба получила подтверждение. Исследования, проведенные с помощью микроэлектродов, показали существование нейронов, которые отвечают только на то или иное свойство стимула, например на направление движения, наклон линии и т. д. Я думаю, что сегодня большинство нейрофизиологов согласны с мнением Хебба о том, что единица восприятия соответствует нейронной единице (рис. VI-7).
Гипотеза, предложенная в первой части этой главы, говорит о том, что выбор, сделанный Хеббом, был единственно возможным и что возникновение структур возбуждения должно зависеть, конечно, от возбуждения специфических клеток, но что эти динамические структуры становятся в какой-то степени независимыми от клеточных единиц, так как определяются анатомией соединений, синаптическими и дендритными микроструктурами мозга. При соответствующих условиях эти узоры, образуемые с помощью синаптических систем, служат неврологическими эквивалентами единиц восприятия. Я согласен с предположением Хебба о том, что то, что мы осознаем, во многом зависит от прошлого опыта организма, но я буду отличать это явление от того непосредственного психологического настоящего, принимающего форму Образов, на котором основана предлагаемая гештальтпсихологией целостная концепция восприятия. Хебб смешал историческое развитие процесса узнавания с тем, что составляет содержание Образов. Чтобы составить программу, которая позволит компьютеру произвести вычисления, требуются многие часы работы, сама же машина выполнит эти вычисления за микросекунды. Необходимо много времени, чтобы научиться узнавать незнакомые структуры, но даже младенцы в возрасте нескольких недель (см. гл. VIII)
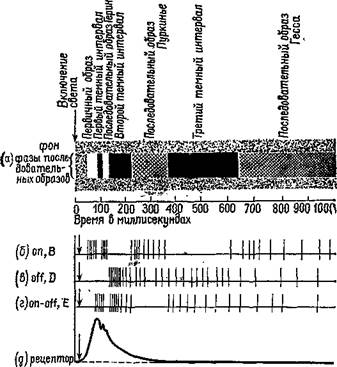
Рис. VI-7. Корреляция субъективных зрительных последовательных образов человека и разрядов нейронов в сетчатке и зрительной коре кошки после кратковременной вспышки света (300 лк) (взято из: Grusser and Grutzner, 1958, с изменениями): а — схс^ма сменяющих друг друга фаз последовательных образов (по: Froh-lich, 1929). Время в миллисекундах, б-г — схематическое изображение ответов on, off и on — off нейронов сетчатки и различных типов (В, D, Е) кортикальных нейронов; д — ответ рецептора наружного плексиформного слоя сетчатки при внутриклеточной регистрации. Вспышка света указана стрелкой, направленной вниз. В б-д стрелка сдвинута вправо, потому что в экспериментах Фре-лиха латентный период ощущения на 20—40 мсек длиннее латентного периода ответов у кортикальных нейронов кошки. Затененная область, окружающая последовательные образы, означает фоновое освещение и фон собственного света сетчатки.
В схеме объединены сетчаточные и кортикальные ответы, хотя последние характеризуются более низкой частотой и большей периодичностью разрядов. Пауза в первичной активации кортикальных нейронов В совпадает с начальным разрядом нейронов Е. При продолжительном освещении и то и другое соответствует «черной полосе» Шарпантье (Jung, 1961), а при кратковременной вспышке — вероятно, первому темному интервалу. Когда Грюссер л Грюцнер описали в реакции on — off нейронов паузу между 200 и 450 мсек, они соответствующим образом внесли поправки в свою схему (Jung, 1969).
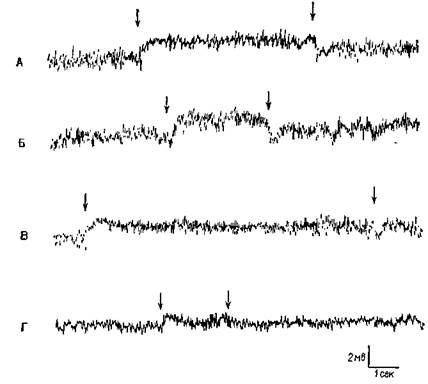
Рис. VI-8. Сдвиг постоянного потенциала (ПП) в слуховой коре в ответ на звуковую стимуляцию. «Активный электрод» — на твердой оболочке мозга над средней эктосильвиевой извилиной; другой электрод — на фронтальной надкостнице. Отклонение вверх означает негативность на твердой оболочке относительно кости. Л — сдвиг ПП в ответ на белый шум; Б — сдвиг ПП в ответ на тон 4000 гц; В — сдвиг ПП, возникающий в ответ на белый шум, возвращается к нулевой линии до прекращения звуковой стимуляции; Г — ответ на звуковые щелчки (50 в сек) и его возвращение к нулевой линии до прекращения стимуляции (Gumnit,
1960).
могут правильно определять относительный размер фигур, находящихся на различном расстоянии (константность величины), и дифференцировать фигуры в различном положении (константность формы). Научение — это только часть проблемы узнавания. Гештальтисты в значительной мере были правы в своем нативизме. Существуют врожденные нервные механизмы формирования Образов, но гештальтисты были неправы, когда утверждали, что это единственный механизм восприятия. Однако сейчас маятник качнулся далеко в другую сторону и существует опасность, что целый ряд интересных явлений выпадет из поля зрения исклю-
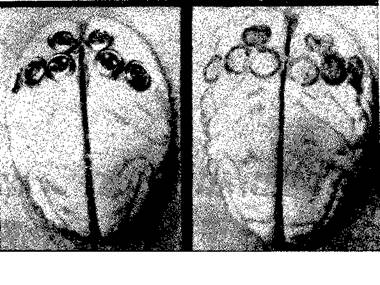
А Б
Рис. VI-9. A — фотография мозга обезьяны со вживленными з него серебряными дисками, содержащими пасту; Б — фотография того же самого мозга после снятия дисков. В данном случав вживлепие было произведено в теменную кору. Другие эксперименты были проведены на затылочной, височной и лобной коре тем же способом или методом множественных точечных инъекций в кору алюминиевой пасты (Stamm
and Warren, 1961).
чительно из-за интереса к проблеме, каким образом мы учимся распознавать.
Когда Хебб писал свою книгу, существовали две основные точки зрения на функционирование нервной системы. Одна из них была хорошо обоснована, другая — нет. Первая касалась вопросов генерации нервных импульсов и их передачи через «инапсы. Другая имела отношение к электрическим полям, создаваемым потенциалами мозга. В. Кёлер построил свою гешталь-тистскую концепцию на основе этих неироэлектрических полей и потерпел неудачу, доказывая их повсеместное существование десять лет спустя после опубликования Хеббом своих взглядов.
Я был счастлив, что смог принять участие в этих исследованиях. Экспериментальное наступление оказалось в целом успешным (КбЫег, 1958), и исследователи в моей лаборатории, а также в других лабораториях, регистрируя и вызывая в мозгу электрические поля постоянного тока, установили -их корреляции с функциями мозга и поведением (Gumnit, 1961; Stamm, 1961; Morrell, 1961). Всякий раз при стимуляции организма через тот или другой сенсорный вход (рис. VI-8) в соответствующих огра-
ничейных областях мозга возникал сдвиг постоянного потенциала. Прямое раздражение мозга постоянным током могло задержать или ускорить научение в зависимости от полярности прикладываемого потенциала. Но я не согласился с Кёлером, когда он стал настаивать на связи между электрическими полями постоянного тока и восприятием. Позднее, когда я закончил эксперименты, в которых я накладывал на поверхность коры мозга алюминиевую пасту, мы снова смогли прийти с ним к согласию. Эксперименты показали, что различение структуры сигналов не нарушается, не--смотря на заметные изменения постоянного потенциала и ЭЭГ (рис. VI-9, VI-10). Кёлер никогда не признавал экспериментов, проведенных Лешли (Lashley, Chow and Semmes, 1951), в которых для опровержения его теории и разрушения нейроэлектри-ческих полей использовалась золотая фольга. Не признавал он также и экспериментов Сперри с перекрестом нервных волокон
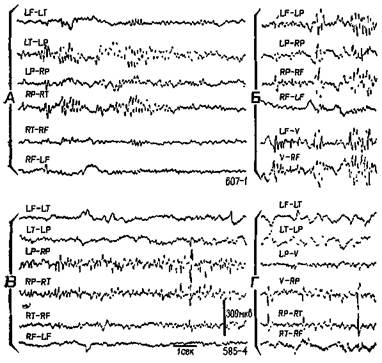
Тис. VI-10. Образец записи ЭЭГ у двух обезьян со вживленными дисками, как показано на рис. VI-9. А и Б — участки записи ЭЭГ до операции, В и Г — спустя 5 месяцев после вживления дисков •с алюминиевой пастой. Биполярное отведение: L — левое, R — правое полушарие, F — лобная кора, Т — височная, Р — задняя темен-лая, V — макушка. Калибровка, как показано (Stamm and Warren.
1961).
(Sperry, Miner and Meyers, 1955), в которых использовались изолирующие полоски слюды. Но когда он познакомился с результатами эксперимента со вживлением дисков с алюминиевой пастой, он воскликнул: «Это опровергает не только мою теорию поля постоянного тока, но и всякую другую современную неврологическую теорию восприятия!»
Позвольте мне коротко остановиться на тех данных, которые накопились в ходе этой дискуссии, чтобы рассеять такой мрачный взгляд на неврологию восприятия. Как было подробно показано в первой части книги, нервный импульс не является единственной важной электрической характеристикой нервной ткани. Другой характеристикой является микроструктура медленных потенциалов. Хотя последние близки к кёлеровским полям постоянного тока, они существенно отличаются от них тем, что они не диффузны, а строго локализованы на соединениях между нейронами или на дендритах и могут даже иметь вид миниатюрных спайков, которые чаще всего при распространении ослабевают. Проведение нервного импульса в любом месте нервной системы ведет к образованию на соединениях микроструктуры медленных потенциалов. Когда нервные импульсы достигают синапсов, генерируются постсинаптические потенциалы. Они никогда не бывают единичными, а образуют динамические структуры афферентных воздействий. Когда постсинаптические потенциалы возникают в дендритных полях мозга, они часто оказываются недостаточно большими, чтобы немедленно вызвать разряд нервного импульса. Поэтому динамические структуры постсинаптических потенциалов образуют узор, который имеет сходство с фронтом волны. Но-этот рисунок медленных потенциалов не является каким-то таинственным фоном, миражем, наложенным на известную нервную функцию. Он представляет собой микроструктуру, состоящую из классических нейронных медленных потенциалов, возникающих в результате прихода нервных импульсов, ожидающих выхода через аксон.
Таким образом, структуры, образуемые на входе и выходе и принявшие форму микроструктуры медленных потенциалов, становятся источником третьей силы в нервной клетке. Все это говорит против концепции «плавающего» поля как возможного нейрофизиологического механизма, обусловливающего целостный характер восприятия. Потребность в этом третьем источнике была обнаружена до его открытия. Лешли глубоко волновала следующая проблема:
«Здесь мы сталкиваемся с такой дилеммой. Нервные импульсы проводятся по определенным, строго ограниченным путям, по сенсорным и моторным нервам и в центральной нервной системе от клетки к клетке и через-определенные межклеточные связи. Однако кажется, что все поведение детерминировано массой возбуждения, формой, отношением или соотношением возбуждения внутри общих полей активности безотносительно к отдельным нервным клеткам. Важна динамическая структура возбуждения.
я не отдельный элемент. Какой тип нервной организации способен отвечать на такую структуру возбуждения при отсутствии специфических, ограниченных путей, обеспечивающих его проведение? Эта проблема имеет отношение к деятельности всей нервной системы, и необходимо сформулировать какую-нибудь гипотезу, чтобы дать направление дальнейшему исследованию» (Lashley, 1942, стр. 306).
Таким образом, Лешли предположил, что модель такой динамической структуры, возникающей в результате интерференции, была бы более адекватным объяснением этого феномена, чем любая из противоположных точек зрения. Однако он не имел ясного представления о такой модели, о том, каким образом может работать этот механизм. Он никогда не говорил о том, что «волны», генерируемые в результате прихода нервных импульсов, образованы хорошо известными классическими нейрофизиологическими процессами: синаптическими и дендритными потенциалами. Таким образом, он никогда не приводил аргументов в пользу существования микроструктуры соединений, в какой-то мере независимой от проведения нервных импульсов, то есть идеи, которая развивается на страницах данной книги. Лешли слишком связывал волновые формы с циркуляцией возбуждения по нейронам, которую он считал недостаточной, и в то же время разграничивал их, когда нужно было объяснять гибкость приспособления. Таким образом, он совсем не развил своей догадки. А ведь даже самая проницательная догадка, чтобы стать очевидной, нуждается в доказательстве. Прервем же на время наше изложение и приведем цитату Лешли, которая дает возможность проследить за ходом его мысли.
«Анатомические работы Лоренте де Но открыли в коре систему перекрестных связей, которые позволяют возбуждению распространяться по поверхности коры в любом направлении. Многие соседние нейроны способны к взаимному возбуждению, и вся система организована как сеть с петлями различной длины и сложности. Последние способны передавать импульсы от одной клетки коры к другой, или повторно возбуждать те точки мозга, которые первыми испытали воздействие стимуляции, посредством систем с возвратной циркуляцией с различными характеристиками.
Заключение о функциональных свойствах такой структуры организации может быть сделано с известной уверенностью. Возбуждение, возникающее в какой-либо точке, должно распространяться от нее по всей системе, поскольку угашение (возникающее в результате активации колец реверберации) будет возникать только после прохождения первоначального импульса. Если бы все системы были однородными, то возбуждение продуцировало бы серию радиально бегущих волн, так как первые волны возбуждения сопровождались бы волной угашения, за которой снова следовало бы возбуждение либо от последующих залпов, прибывающих по сенсорным путям, либо вследствие повторного возбуждения, возникающего на дальней границе зоны угашения. Длительность волн была бы одинаковой, ибо она» зависит от скорости проведения и рефрактерных периодов элементов всей системы. При наличии нескольких или многих точек возбуждения будут возникать интерферирующие динамические структуры.
Чтобы представить себе более простую картину, отвлечемся на время! от тех явлений, которые возникают в результате влияний от колец обратной связи; в этом случае происходило бы нечто аналогичное явлению распространения волн по поверхности жидкой среды. В такой системе интерференция волн создает структуру из гребней и впадин, которая характерна-для всякого пространственного распределения источников волнового движения и остается примерно одинаковой на всей поверхности. Можно предположить, что и в самой коре имеет место процесс, сходный в какой-то-степени с формированием структур возбуждения. Пространственное распределение в коре импульсов, пришедших к ней из сетчатки, не воспроизводит структуру возбуждения сетчатки, но создает условия для появления другой и специфической структуры стоячих волн, которая повторяется по» всей функционирующей области. Непосредственным препятствием этому является то, что возбуждение одной части поля может создавать в другой части состояние рефрактерности для приходящих от остальных частей-мозга импульсов и таким образом блокировать формирование единой динамической структуры. Однако если транскортикальные пути колец реверберации имеют случайную длину, как это, видимо, и есть на самом деле,. то в любой области они не все будут приходить в рефрактерное состояние одновременно и блокирования не будет» (Lashley, 1942, р. 313—314).
В последующих главах мы рассмотрим наиболее современную-и обоснованную точку зрения на «третью силу» в нейрофизиологии восприятия, представление о которой основано на признании существования двух типов микроструктур: нейронной микроструктуры и микроструктуры соединений.
РЕЗЮМЕ
Первые бихевиористы решительно отказывались от изучения субъективного опыта. В результате этого произошло значительное ограничение предмета психологии. Концепция двухпроцесс-ного механизма функционирования мозга устраняет это ограничение, вооружая психологов объективными методами и понятийным аппаратом механизмов соединений, с помощью которых можно изучать перцептивные образы, а также эмоции и мотивы.
Г л а в а VII
ПРИЗНАКИ








