 2015-10-16
2015-10-16 445
445Как мы уже говорили, наблюдения Бернштейна показывают, что единство действия может быть достигнуто независимо от частных видов движения или количества сокращений каких-либо частных мышц или групп мышц. Действие может осуществляться лишь потому, что движения согласуются с «полем внешних сил», вовлеченных в действие. Как мы уже отмечали при рассмотрении гамма-эфферентной сервосистемы, существенные изменения в этой системе компенсируются указанным механизмом, так что постоянно сохраняется состояние готовности к движению. Таким образом, стабильность состояния достигается за счет специальных приспособлений и компенсаций внешних сил, а представительство в коре больших полушарий этого механизма включает эти параметры. Фактически центральный нервный механизм и состоит в основном, из этих приспособлений и компенсаций; следовательно, центральное представительство должно быть организовано таким образом, чтобы быть «зеркальным отражением» поля внешних сил (аналогичный вывод был сделан и в гл. XII).
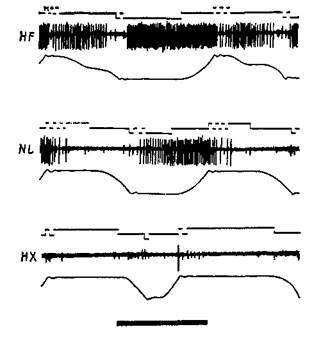
Явное доказательство того, что нейроны моторной коры — особенно те, от которых идут пути на периферию, — действительно чувствительны к воздействиям, вызывающим мышечные сокращения, было получено в серии других остроумных опытов. В этих экспериментах Э. Эвартс (1967, рис. ХШ-6) обучал обезьян работать рычагами, к которым были подвешены различные грузы, чтобы затруднить движения. Когда обезьяны научались двигать рычаг, в момент выполнения задания регистрировалась активность нейронов, расположенных в моторной коре. Опыты показали, что эти нейроны активируются до внешних признаков начала движения (по миографической записи). Тщательный анализ установил, что эта электрическая активность зависит от величины необходимых усилий, а не от величины смещения рычага. Более того, нейроны реагировали главным образом на изменение требуемой силы, хотя не исключено, что некоторая активность связана и с величиной необходимого усилия (рис. ХШ-7).
Тот факт, что моторная кора предвосхищает параметры усилия, очень важен. Поскольку рефлексы организуются
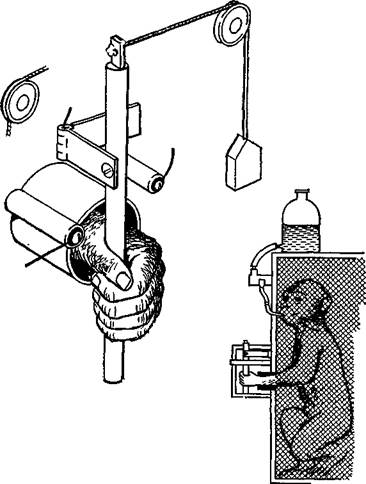
Рис. XIII-6. Аппарат для изучения поведения обезьяны с одновременной регистрацией моторного эффекта в клетках прецент-ральной коры. Левая рука обезьяны просовывается через отверстие в прозрачной перегородке, прикрепленной к передней стенке клетки. Чтобы получить фруктовый сок в качестве вознаграждения, обезьяна должна схватить вертикальный стержень, присоединенный к шарниру, и передвигать его взад и вперед между двумя упорами. Чтобы сработал соленоидный приемниь-, который «выдает» фруктовый сок, интервал между соприкосновениями стержня с контактами должен составлять 400—700 мсек и предварительное движение в другом направлении также должно совершаться в это же время (Evarts, 1968; см. Милнер, 1973).
1сек
Рис. XII1-7. Активность клетки моторной коры обезьяны в условиях, изображенных на рис. XIII-6. Показана активность клетки при трех видах нагрузки. Использовались следующие три вида нагрузки: 400 г на сгибатели (верхняя запись), 400 г на разгибатели (нижняя запись) и без нагрузки (средняя запись). Видно, что активность нейрона значительно возрастает при нагрузке на сгибатели, то есть при увеличении силы сгибания. Напротив, клетка почти полностью «замолкает» в те периоды, когда движение совершается при нагрузке на разгибатели (Evarts, 1967).
сервомеханизмами, центральное представительство этих рефлексов составляет не запись длины или напряжения мышц, а запись параметров приспособлений и компенсаций, соответствующих тем изменениям внешних сил, которые вовлечены в движение. Вследствие конвергенции этих процессов в центральном представительстве движений формируется образ не только прежних и текущих изменений внешних сил, воздействующих на систему, но — благодаря быстродействующим расчетам движений, совершающимся в мозжечке, — и тех изменений, которые должны произойти при продолжении двигательной активности.
Таким образом, моторную кору можно рассматривать как сенсорную по отношению к действию. Она участвует в пространственной модуляции состояний готовности благодаря своим связям
с базальными ганглиями и с механизмом быстродействующего-расчета параметров конечного эффекта действия, осуществляющегося при участии замкнутой мозжечковой системы. Формирование Образа результата зависит — как и всюду в коре — от того, что мы называем «нейронным топографическим представительством», то есть «от в высшей степени избирательного протекания активности в горизонтально расположенных корковых сетях, которые возбуждают и тормозят кортикофугальные нейроны» (Phillips, 1965). То, что это моторное представительство действительно во многих отношениях сходно с тем, которое формируется в сенсорных зонах мозговой коры, может быть установлено при исследовании рецептивных полей нейронов. Результаты этих экспериментов (Welt et al., 1967, табл. ХШ-1) показали, что «сенсорная конвергенция в моторной (или сенсорной) коре накладывается на топографически сходные выходные структуры в виде радиальных рядов, диаметр которых, как оказалось, равен 0,1—0,4 мм. Таким образом, нейроны с фиксированными локальными рецептивными полями образуют радиально ориентированную структуру (референтную систему) по отношению к одним и тем же периферическим выходам... Вперемежку с этими клетками, имеющими локальные поля и составляющими 3Д общей массы клеток, расположены нейроны с широкими, вертикально ориентированными или лабильными полями, которые перекрывают локальные поля» (стр. 285). Здесь, в моторной коре^ обнаружены явления, которые еще надо найти в зрительной коре '.
Остается сделать еще только один шаг в анализе работы моторной коры, а именно указать на то, что уже прекрасно выполнено Родиеком в отношении сетчатки: выяснить, какая именно трансформация точно отражает взаимоотношения между корой и периферией.
Итак, контроль над поведением со стороны нервной системы достигается главным образом посредством воздействия на рецеп-торные функции. На рефлекторном уровне чувствительность рецепторов к грузу возбуждает и направляет приспособительный контрпроцесс в сервомеханизме. Сумма таких приспособительных процессов и составляет фоновое тонически устойчивое состояние, на котором возникают новые приспособительные действия. Приспособления широкого диапазона, например изменения позы, контролируются экстрапирамидной системой мозга, в которую входят базальные ганглии и передние структуры мозжечка, тогда как более дискретные движения, такие, как печатание на пишущей машинке, как игра на фортепиано, регулируются быстродействующей неомозжечковой системой,.
1 Теперь эти факты уже получены (Spinelli, Pribram and Bridgman, 1971).
Таблица XJ1I-1
Типы рецептивных колей и антидромные латентные периоды реакций клеток моторной коры
(Из- «Neurophysiologtcal Basic of Normal and Abnormal Motor Activities», Ed, Фу M. D. Yahr and D P. Purpura, 1967, New York.)
| Антидромные | Антидромные латентные | |||
| Типы рецептивных | латентные | Адекватные | периоды | Коли- |
| полей | периоды в мсек. Средняялв.а. | стимулы | подгрупп в мсек. Средняя±з.(1. | чество клеток |
| -Фиксированный | 1,99 ±1,03 | Поглаживание по шерсти | 2,57 ±1,08 | |
| Локальный | S. Е. = 0,17 | Прикосновение | 1,88 ±1,07 | |
| (N = 40) | Давление | 1,72 ±0,92 | ||
| Комбинированное воздействие | 1,69 ±0,65 | |||
| "Фиксированный | 1,94±1,19 | Поглаживание по шерсти | 1,45 ±0,93 | |
| Широкий | S. Е. = 0,22 | Прикосновение | 1,94 ±1,24 | |
| (N = 30) | Давление | 2,75 ±0,77 | ||
| Комбинированное воздействие | 3,13±1,15 | |||
| Лабильный | 1,45 ±0,59 | Поглаживание по шерсти | 1.40 ±0,42 | |
| S.E. = 0,16 | Прикосновение | 1,48 ±0,38 | ||
| (N = 18) | Давление | 1,58 ±1,01 | ||
| Комбинированное воздействие | 1,27 ±0,34 |
Сравнивались в простом рандомизированном плане антидромные латентные периоды реакции 88 корковых клеток, сгруппированных по типам рецептивных полей -Ответы группы «лабильных» клеток отличались от ответов обеих групп <Фикси1 рованных» клеток более чем на 1% not—критерию в предположении равенства групповых дисперсии. v*
s. d.— стандартное отклонение
S. Е.— средняя ошибка
См. описание сходства роли лабильных клеток и чувствительных клеток узко-яаправленного действия в корковых структурах Вернера (рис, VH-7), основанное/на -концепции т—о—Т—Е (Welt et. al. 1967).
•осуществляющей экстраполяционные расчеты. Точный механизм этого центрального контроля за движениями пока еще не изучен, -однако то, что мы знаем, достаточно, чтобы утверждать, что в этом процессе участвуют явления моделирования периферических сервомеханизмов и что такое моделирование осуществляется посредством механизмов коррекции. Наконец, предлагаемая концепция радикально изменяет наши представления о функциях прецентральной моторной коры. Эта часть мозга, как было показано, является сенсорной по отношению к действиям. Образ конечного результата формируется и непрерывно -мгновенно видоизменяется благодаря нейронным голографиче-
ским процессам, так же как и перцептивный Образ. Однако Образ ожидаемого результата создается на основе уже известных из прошлого опыта антиципации тех внешних сил и их изменений, которые требуются для выполнения задания. Эти поля сил,, оказывающие влияние на мышечные рецепторы, становятся параметрами сервомеханизма и непосредственно (через таламус) или опосредствованно (через базальные ганглии и мозжечок) связываются с моторной корой, где они сопоставляются с быстродействующими расчетами мозжечка по предвосхищению результата и последующего звена действия. Когда ход действия становится предсказуем на основании тенденций предшествующих успешных предсказаний, может быть сформирован Образ конечного результата для управления окончательной фазой двигательной активности.
Эта модель, которая вытекает из результатов приведенных исследований, существенно отличается от представлений о моторной коре как о клавиатуре, к которой стекается активность остальной части мозга, чтобы разыгрывать мелодию движения. Эта модель, точнее, представление о том, как совершить движение, кодируется в моторной коре и превращается в эффективные действия, акты — внешние выражения мозговых процессов.
В следующих главах мы рассмотрим вопрос о том, как эти внешние представительства движений модифицируются в последовательности действий. Последующая часть книги посвящен» исследованию действий, связанных с процессом коммуникации.
РЕЗЮМЕ
Из ряда мозговых процессов, участвующих в организации поведения, один привлекает особое внимание. Этот процесс включает в себя действие моторного механизма коры больших полушарий и коры мозжечка, формирующего Образ конечного результата с помощью быстродействующих расчетов. Посредством этого-механизма осуществляется экстраполяция последствий непосредственно предшествующих движению изменений во внешнем поле, которые воздействуют на мышечные рецепторы. В этом Образе-конечного результата кодируются, следовательно, внешние явления (например, силы), а не наборы мышечных сокращений. Образ-конечного результата регулирует поведение так же, как термостат регулирует температуру: на диске прибора закодированы пе включение и выключение котла, а лишь пределы допустимо»' температуры.
Глава XIV
КОМПЕТЕНЦИЯ
БИХЕВИОРИЗМ: СТИМУЛЫ, ОТВЕТЫ И ПОДКРЕПЛЕНИЕ
В предыдущих главах мы проводили различие между движением и действием. Движения, то есть организованные мышечные -сокращения, создают набор поз, двигательный контекст или матрицы, на основе которых формируются акты поведения. В нервный механизм, контролирующий действие, включена информация, поступающая из внешней среды, которая создает нечто вроде силовых полей, но в ней не содержатся сведения о длине той или другой мышцы и о расположении суставов. Таким обра--зом, любое направленное действие может выполняться относительно широким набором потенциальных движений. Перефрази-■руя утверждение Гранита относительно рефлексов (на которое мы ссылались в гл. XII), можно сказать, что основными проблемами поведения являются не проблемы движений, как таковых, хотя поведение осуществляется посредством движений. Основные проблемы связаны с процессами настройки, или установки, различных сервомеханизмов, посредством которых вводятся в действие мотонейроны. Эти процессы настройки, как правило, включаются в действие под влиянием факторов внешней среды, действующих в данной ситуации.
Неожиданно оказалось, что вопрос, поставленный нейрофизиологом, входит в сферу интересов психолога. Специфика этих внешних условий, определяющих действие, и есть то, что всегда изучает бихевиорист, пытающийся построить свою концепцию на анализе организации внешней среды, значимые связи которой получают объяснение через поведение организма. Бихевиористы правильно считают, что стимулы, ответы и подкрепление получают свое значение на основании внешних, а не внутренних условий существования организма, хотя необходимо учитывать и поведение организма, без которого картина останется неполной. Почему тогда психологам не ограничиться только «изучением доведения»?
Отношения между стимулом, ответом и подкреплением сложнее, чем это представляется бихевиористу. Хотя центр интересов бихевиориста меняется, все бихевиористы — за небольшим исключением — представляют себе эту связь как прямую последовательность. Стимул -> ответ -*- подкрепление является той самой последовательностью, которая видна наблюдателю. Однако то, что эта очевидная последовательность есть результат более сложных взаимоотношений, стало ясно только тогда, когда обнаружилось, что мозг обеспечивает этот порядок.
Возьмем, например, тот факт, что рефлекс организуется не па схеме дуги S -*- R, a с помощью сервосистемы. Этот факт оказал важное влияние на понимание сущности стимула и ответа. Существование механизмов обратных и предваряющих связей усложняет всем известные представления Ньютона и Шерринг-тона о линейной связи стимула и реакции. В результате перед бихевиористами возникли две возможности: либо игнорировать внутреннюю сложность этой системы, либо пытаться объяснить ее и дать трактовку тем изменениям, которые претерпевает эта система. В большинстве случаев бихевиористы игнорировали сложность системы. Однако они не могут уйти от того, что стимулы сами находятся под воздействием возникающих ответов иг напротив, что ответ можно полностью объяснить, только учитывая вызвавшие его условия. Иначе говоря, с точки зрения бихевиоризма стимул и ответ взаимно предполагают друг друга. Эта дилемма может быть разрешена только в том случае, если признать взаимосвязь между S и R. Схема S з=ь R имеет отношение не только к неврологии, но и к логике. Математический подход раскрывает глубокое значение этого взаимодействия (Estesv 1959). Предполагается, что стимулы являются элементами этой системы и что их отдельные составляющие соответствуют ответам (рис. XIV-1). Можно сказать, что стимулы — это систематизированные объекты, а ответы — процесс классификации этих стимулов. Однако решение этой дилеммы (S — R) предъявляет очень высокие требования к тому, как освещать факты. В современной психологии большинство расхождений в формулировках происходит из-за неспособности до конца понять эти реципрокные взаимоотношения между стимулом и ответом. В физиологической психологии основные противоречия возникают между теми, кто описывает полученные данные на языке стимулов, и теми, кто предпочитает язык ответов. И конечно, противоречия часто происходят из-за смешения этих языков.
Согласно такому логически и неврологически обоснованному пониманию взаимоотношений между S и R при описании фактов следует придерживаться либо языка стимулов, либо языка ответов, но не обоих сразу. Противоречия возникают потому, что слова, используемые каждым языком, те же самые, хотя их смысл совершенно различен. Большинство отраслей психологии
взбежало 9tofo противоречия, ограничиваясь лишь тем или другим типом анализа. Так, психологи, занимающиеся восприятием, и психофизики пользуются при описании полученных данных.языком стимулов: механизмы ответов выводятся из организующих свойств стимулов. Напротив, психологи, придерживающиеся
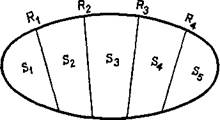
Тис. XIV-1. Концепция, согласно которой реальность описывается в терминах стимулов (5.) и ответов (Лг). Каждому ответу соответствует
свой стимул.
концепции оперантного обусловливания, и сторонники психологии познавательных процессов пользуются языком ответов: стимулы определяются теми их свойствами, которые вызывают ответ. Таким образом, возникают две дополняющие друг друга ^объяснительные схемы, и физиологические данные иногда относятся то к одной, то к другой из них (рис. XIV-2, 3). Путаница возникает тогда, когда схема рассуждения четко не определена. Исходя из этого, рассматривая во второй части книги субъективный опыт, мы придерживаемся языка стимулов, принятого у психофизиков, занимающихся сенсорными процессами, и психологов, исследующих проблемы восприятия. В третьей части мы воспользуемся языком ответов, принятым у психологов, занимающихся оперантным обусловливанием и познавательными процессами, так как этот язык больше соответствует анализу поведения.
Язык стимулов Язык нейрофкзиологии Язык ответов
Психофизика Павловская теория ус- Теория оперантного
ловных рефлексов обусловливания
.Гештальтпсихология Психология Хебба Познавательная психо-
логия
Рис. XIV-2. Пример используемых в психологии «языков»: язык стимулов, язык ответов и нейрофизиологический язык.
Возможно, наиболее интересным для анализа этой проблемы «позиций бихевиоризма является объективность, достигнутая в определении подкрепления, когда схема S-> R, или концепция ^флекторной дуги, заменяется теорией сервомеханизмов. Аген-
том, запускающим сервомеханизм, является проба на соответствие-или несоответствие между внешними образами, текущим «состоянием» и характером энергии, воздействующей на это состояние. Тйким образом, то, что психологи называют ближайшим («проксимальным») стимулом, зависит от состояния системы, открытой" для ввода информации, то есть от «установок» и «ожиданий» — по терминологии психологии познавательных процессов. Другие рассуждения ставят подкрепляющие свойства среды в зависимость от совпадения условий, которые вызывают данное-поведение, с теми условиями, которые оно само вызывает. В этой главе, а также в гл. XV и XVI излагаются различные данные нейрофизиологии и нейропсихологии, полученные при изучени» процессов подкрепления — механизма, посредством которого осуществляется регуляция поведения. Теперь мы должны приступить к анализу организации процесса поведения во времени.








