 2015-10-13
2015-10-13 1152
1152Чтобы получить достоверные результаты о состоянии ресурсов животных, экологической и территориальной структуре их популяций необходимо уметь различать, а потом фиксировать все следы жизнедеятельности разных видов зверей и птиц. Особенно важны такие знания при учете численности наземных видов животных, в частности при зимнем маршрутном учете (ЗМУ), основанном на подсчете пересечений отпечатков следов на учетном маршруте. Анализ многолетних результатов ЗМУ в различных регионах России показал существенное занижение итоговых оценок в сравнении с фактической численностью. Мы считаем, что определяющей ошибкой в ЗМУ является игнорирование динамики следовой активности животных, меняющейся в связи со сменой климатических параметров среды, обеспеченностью доступными животных кормами, физиологическими перестройками в организме и другими факторами.
Многолетние исследования зимнего поведения охотничьих животных в разных регионах России учеными ВНИИОЗа (Корытин, 2009) показали, что следовая активность зверей (показатель учета или среднесуточное число следов на 10 км маршрута) одного вида в одном и том же урочище может иметь по дням снежного периода колоссальную разницу: по грызунам она может достигать ста и более (244!) раз, по хищникам и копытным максимальные различия исчисляются десятками раз (в пределах сотни).
Установлено наличие резких колебаний количества следов на одном и том же маршруте при ежедневных учетах. Обнаруженная колоссальная изменчивость данного показателя характерна для очень многих наблюдавшихся видов, что указывает на то, что разовые маршрутные учёты на одном месте не могут адекватно отражать численность зверей. Лишь многократно повторяющиеся маршрутные учеты дают возможность в каком-то приближении правильно характеризовать численность (Корытин, 1974).
Динамика изменений следовой активности по дням у зверей разных видов на одном и том же постоянном маршруте иногда имеет сходство, но изменения происходят независимо друг от друга (Корытин, 2009). В изменениях следовой активности по дням у зверей какого-либо вида может возникать определенная ритмичность, но она не постоянна и сменяется аритмичными колебаниями. Хищникам свойственна более равномерная следовая активность, чем животным из разряда жертв.
Динамика следовой активности по месяцам снежного периода у многих изучавшихся животных весьма изменчива, колеблется по годам, однако носит печать видовой специфичности. Например, в Кировской области для зайца-беляка при наличии вариаций более характерно постепенное снижение числа следов вплоть до февраля, а в марте - скачок вверх; для белки - обилие следов по осени, снижение их количества зимой (в декабре, январе) и возрастание к весне (в феврале и особенно в марте); для лисицы - повышение числа следов в зимние месяцы (в декабре, январе, феврале) и более низкий уровень осенью и весной (в ноябре и марте); у ласки также имеет место усиление следовой активности в серединные месяцы снежного периода. Для зверей, у которых спаривание происходит в снежный период, максимальное количество следов обычно встречается в тот месяц, когда происходит гон.
Ритмика следовой активности по суткам некоторых животных, например белки, на территориях, удаленных друг от друга (даже на расстояние 800 км) имеет сходство. Сходство обнаруживается для большего числа видов. Разница в активности обусловлена различиями в условиях существования. Повышение двигательной активности животных, возможно, обусловлено увеличением плотности населения людей.
В связи с явлением сходства ритмики подвижности дальних соседей относительные маршрутные учеты в разных районах лучше проводить в одни и те же дни, и вести математическую обработку учетных данных, группируя их для сравнения по конкретным датам.
Реакция на лыжню, отвороты от нее присущи не только зверям, которых принято считать осторожными. Процент отворотов варьирует, но для каждого вида характерен свой количественный уровень. Например, соболю, шедшему шагом, а не прыжками, присуще усиленное мечение лыжни мочой и экскрементами (без отворотов от лыжни).
По месяцам снежного периода число отворотов варьирует, но для всех изучавшихся видов характерно возрастание их числа с осени к середине зимы - январю, а потом вплоть до марта - снижение. У хищников кривая более полога, чем у грызунов. У тех и других она коррелирует с температурой воздуха.
Реакция на лыжню зависит от периода суток. Более всего звери боятся лыжни в вечерний период, меньше - ночью, еще меньше утром и менее всего днем. Процент отворотов коррелирует со следовой активностью в течение суток: чем больше следовая активность, тем больше процент отворотов.
Играет роль рекреационный фактор: звери Вятских лесов пугаются лыжни чаще, чем в Подмосковье. Звери относятся к лыжне смелее в своих коренных, наиболее свойственных виду местообитаниях. Неравномерное расположение отворотов на маршруте указывает на влияние дополнительных, кроме самой лыжни, факторов.
Для нейтрализации влияния лыжни на поведение животных и результаты учетов, для повышения их достоверности предлагается внедрение учетной полосы (до 5 м справа и слева от лыжни), перекрывающей поле эмоционального воздействия лыжни на животное.
Переходы. Для лисицы характерно наличие регулярно из года в год использование переходов. Имеются также постоянные, но иногда угасающие переходы: ими лисицы ходят не каждый год. Для горностая свойственно переложное использование территории. Обычно ни один из интенсивно используемых переходов горностая в какой-либо зимний сезон на будущий год не сохранялся. Пространственное размещение горностая менялось по годам, но за ряд лет вся территория была использована полностью. Если тот или иной участок эксплуатируется горностаем интенсивно, то на следующий год он обычно пустует или на нем встречается минимальное количество следов. Ласка в меньшей степени, чем горностай, приурочена к определенным местам.
Интенсивное использование по годам какого-либо участка местности зайцем-беляком постепенно возрастает, а затем постепенно падает, вероятно, в связи с истощением кормовой базы.
Имеются места, где следы животных из года в год встречаются в большем количестве, чем по их окрестностям. Эти «общие переходы», «постоянные дороги» зверей не абсолютно постоянны, время от времени забрасываются, а затем снова возобновляются.
Видовой состав зверей, пользующихся общим переходом, не стабилен, меняется по годам. В пределах каждой стации находится участок, наиболее предпочитаемый животными для переходов.
Скопление следов по разным стациям видоспецифично. Характерное распределение следов на местности у разных зверей в зависимости от их численности меняется по-разному. Если при возрастании численности лося и ласки площадь, занятая их следами, увеличивалась, то для белки и мышевидных грызунов наблюдалось обратное явление: при высокой численности общая площадь, занятая следами этих зверьков, уменьшалась, но росла их плотность на определенных, вероятно, наиболее кормных участках.
Для зайца-беляка, лисицы при низкой относительной численности характерно наличие участков, используемых для переходов более интенсивно; при высокой относительной численности таких мест встречалось меньше. В распределении следов на местности хищников и жертв имеется некоторое сходство, но оно незначительно.
Картина пространственного размещения следов, а следовательно, и использования зверями территории, меняется, пульсирует. Среди изучавшихся видов наиболее стабильно расположение следов в пространстве у лисицы, белки и мышевидных, более изменчиво у мелких куньих - ласки и особенно горностая, а также у зайца-беляка.
Метод постоянного маршрута со значительным количеством постоянных ориентиров дает возможность изучать пространственное распределение животных, зависимость его от стаций и других факторов, а также учитывать расположение переходов зверей с большой точностью.
Следовая активность в окладах. Выявление всех зверей на учетных площадках не простое дело. Учет численности животных путем оклада с последующим прогоном трудоемок и сложен в организационном плане, поскольку требует участия нескольких человек. Чтобы избежать прогона Я.С. Русанов (1961) предложил двукратный маршрутно-окладной метод, сущность которого заключалась в трехкратном прохождении вокруг учетной площадки: при первом осуществляется затирание следов, при втором и третьем - учет следов животных, вышедших с площадки. В основу этого метода положено предположение, что за два дня учета находящиеся в окладе животные проявят свое присутствие следом.
Сколько времени звери таятся в окладе. В Белоруссии был применен «трехкратный маршрутно-окладный метод учета» (Козло, 1973), при котором учет проводился три дня подряд. «В результате значительно повышается точность учета», - пишет П.Г. Козло (1973), - и особенно тех видов, которые в силу присущих им поведенческих особенностей или под влиянием внешних факторов среды находятся внутри лесного квартала и не дают следа по просекам более одного-двух дней».
Многодневные маршрутно-окладные учеты ВНИИОЗ под руководством С.А. Корытина показали, что трехкратный учет также не гарантирует выхода с учетных площадок всех находившихся там особей (табл. 8).
Наблюдения проводились одновременно на двух маршрутах: в Кировской и Московской областях в зимний сезон 1978-1979 г. Каждый маршрут опоясывал по пять площадок площадью по 25 га.
Принимались во внимание лишь выходы зверей, которых, по расчетам, не должно было быть на учетной площадке. Например, по соотношению выходных и входных следов на площадке находятся два лося, а на следующий день при обходе обнаружены выходы из оклада не двух, а трех лосей. Очевидно, третий лось раньше находился в окладе, то есть учитывались звери, таившиеся на учетной площадке, не дававшие о себе знать следами.
Были получены следующие результаты. Зайцы-беляки в Кировской области обнаруживались не только на третий, но и на шестой и седьмой день наблюдений, а в Подмосковье на 10, 11 и 13 день наблюдений! Белки обнаруживались на 4 и 5 день. Ласки в основном дали о себе знать в первые 4 дня, но один зверек объявился на 10 день. Горностаи обнаруживались вплоть до 8-го дня. Куница лесная вышла с площадки через 4, а лисица - через 3 дня.
Основная масса затаившихся лосей вышла из окладов в первые 5 дней, но некоторые особи выявлялись на 7, 9 и даже 10 день наблюдений. Кабан появился через 3 дня. Таким образом, основная масса таившихся зверей была выявлена в первые 5 дней наблюдений, но были особи, объявляющиеся и на 10 день, а отдельные рекордсмены показали себя лишь на 13-й день.
То есть можно с уверенностью сказать, что трехдневный оклад (а тем более двухдневный) не гарантирует выявления всех особей, находящихся внутри даже таких небольших учетных площадок - в 25 га.
Таблица 8
Динамика выхода из десяти окладов (площадью 25 га) зверей разных видов (Корытин, 2009)
| Вид | Место наблюдений | Дни наблюдений* | ||||||||||||||||
| Заяц-беляк | Кировская обл. | + | + | + | ||||||||||||||
| Московская обл. | + | + | + | + | + | |||||||||||||
| Белка | Кировская обл. | + | + | |||||||||||||||
| Московская обл. | + | + | ||||||||||||||||
| Ласка | Кировская обл. | + | + | + | + | |||||||||||||
| Горностай | Московская обл. | + | + | + | + | + | + | + | ||||||||||
| Куница | Московская обл. | + | + | + | ||||||||||||||
| Лисица | Московская обл. | + | ||||||||||||||||
| Лось | Кировская обл. | + | + | + | + | + | + | + | ||||||||||
| Московская обл. | + | + | + | + | ||||||||||||||
| Кабан | Московская обл. | + | + |
Примечание: -* обозначены дни, когда из оклада выходили особи, бывшие там к началу наблюдений.
Несомненно, двух- и трехдневные учеты дают заниженные показатели численности животных. Судя по наблюдениям сотрудников ВНИИОЗа, значительно точнее пятидневные учеты (не считая дня затирания следов), хотя они в ряде случаев также не обеспечивают абсолютной точности.
Динамика откочевок и подкочевок зверей на малых территориях. Многодневные подсчеты следов по периметру учетных площадок дают возможность проследить динамику пребывания зверей внутри окладов. Кривые вычерчивались, начиная с цифры, показывающей исходное количество зверей в окладе. Она определялась в конце периода наблюдений с учетом всех случаев ухода животных из оклада. Например, в первый день учета на площадке отмечено пять входных следов лисицы и три выходных. Следовательно, в окладе оставалось два зверя. На второй день учета обнаружилось пять выходных следов и один входной. Следовательно, к началу учета на площадке находилось не менее четырех зверей, а сейчас имеется один. И так далее, пока звери, находившиеся на учетной площадке, к моменту начала учетных работ, не обнаружат себя следами.
Количество зайцев на учетной площадке в Подмосковье (зимний сезон 1979 г.) в первые 14 дней наблюдений при наличии колебаний кривой постепенно нарастало с 26 особей до 68, а затем в течение 17 дней численность этих зверьков постепенно падала за счет откочевки зверьков и достигла к 31 дню наблюдений почти исходного уровня - 35 штук. Не исключено, что фактически число зайцев на площадке к концу наблюдений было меньше указанного, т.к. часть особей могла погибнуть внутри площадки от зубов хищника или естественной смертью. Чем было вызвано это скопление зайцев (и увеличение их плотности до одной особы на 1,8 га) - неясно. Возможно, наличием изобильных кормов или необходимостью внутривидовых контактов. Может быть, у млекопитающих существуют свои «конференции»? Ведь наблюдаем же мы иногда временные многочисленные сборища сорок, ворон и других птиц.
Скопление зайцев не осталось незамеченным для лисиц, численность которых на учетной площадке за этот период возросла с одной особи до 9. Зайцы начали быстро откочевывать с площадки после того, как численность лисиц на ней достигла четырех штук. Затем темп откочевки зайцев несколько снизился, но лисицы продолжали прибывать на место вероятной поживы. Вполне вероятно, что откочевка зайцев была вызвана накоплением на площадке хищников - лисиц, которые распугивали зверьков и производили опустошения в их рядах. Появление микроскоплений зайца-беляка в годы депрессии его численности в Кировской области отмечает и В.А. Агафонов (1988).
Нечто подобное наблюдается весной во время паводка, когда зайцы и другие животные, теснимые водой, скапливаются на островках. Но это скопление происходит вынужденно, а зимнее скопление, наблюдавшееся сотрудниками ВНИИОЗа, происходило на добровольной основе.
На этой же учетной площадке в значительно меньших масштабах происходило скопление белки. Численность ее поднялась с 22 особей до 35. Численность на этом уровне держалась около 8 дней, а затем пошла на убыль, достигнув минимума на 28-ой день наблюдений, - в тот же день, что и численность зайцев. Численность на учетной площадке горностая, как и лисицы, возрастала: с 8 штук до 15 в течение 8 дней, а затем продолжала оставаться на этом уровне до конца наблюдений. Численность третьего хищника - куницы - тоже несколько возросла к концу наблюдений - с 3 до 5 особей.
Численность копытных - кабана и лося - на учетной площадке изменялась в период наблюдений незначительно.
Динамика пребывания зверей в окладе на другой учетной площадке - в Зуевском районе Кировской области (примерно в этот же период) - была иной. Заяц-беляк, поднявшись в численности за первые 9 дней наблюдений с 2 особей до 12, затем в течение 25 дней оставался примерно на том же уровне численности, а в оставшиеся 13 дней возрос в числе до 27 особей.
С белкой другое: первые 27 дней наблюдений она имела численность около 8-14 особей; потом за 7 дней возросла до 30 особей и 9 дней пребывала на учетной площадке примерно в этом количестве. А затем произошла откочевка: в течение 5 дней численность снизилась вдвое - с 28 особей до 13. Нечто подобное наблюдалось с лаской (Корытин, 2009).
Лосей в первые 14 дней наблюдений на учетной площадке было около 15. Потом число их возросло до 21, а затем постепенно снизилось до 9.
Судя по имеющимся материалам, численность на небольших территориях (в пределах 25-125 га) даже таких оседлых видов, как белка и заяц-беляк, может колебаться на протяжении 1-2 месяцев за счет откочевок и подкочевок в значительных пределах, изменяясь до двух и более раз. Интересно, что колебания эти при наличии мелких скачков происходят в основном более или менее постепенно. Такой характер изменений указывает на то, что эти флуктуации численности обусловлены не случайными передвижениями, а вызваны какими-то местными причинами: колебаниями условий среды обитания, а может быть, изменениями, происходящими внутри микропопуляций.
Трудности определения длины суточного следа зверя. Пересчетный коэффициент определяется обычно путем тропления. Это слишком трудоемкий способ, качественное практическое исполнение которого затрудняется большим количеством помех: трудностью в идентификации следа при выходе одного зверя на след другого (что нередко наблюдается, например, при троплении лисицы), наличием нескольких лежек на суточном пути зверя, спугиванием зверя во время тропления и т.д. Измерить точно длину суточного наследа животного - трудное дело. След лисицы, зайца или другого животного зачастую имеет такую извилистую и причудливую форму, что измерить длину всех составляющих его петель не представляется возможным. Тем более, что из-за отсутствия подходящего прибора для измерения расстояния промеры длины наследа производятся шагами, тогда как размеры петель на следах мелких зверьков (например, ласки, горностая) бывают меньше длины парного шага, который является единицей измерения. В некоторых случаях следы на жировках бывают так запутаны, что разобраться в них невозможно, а тем более вытропить. В последнем случае учетчик, проходя на лыжах, вытаптывает площадку со следами раньше, чем промерит и половину всех петель.
Определение длины суточного следа иногда затрудняется уходом зверя под снег или на деревья. Например, куница, увидев лыжню, часто поднимается на деревья и идет верхом; верховой след куницы может заметить далеко не каждый учетчик.
Во время тропления определяется длина суточного следа только одного животного, которого тропят. Чтобы определить длину суточного следа, скажем, восьми видов животных, обитающих в данной местности, нужно сделать минимум восемь троплений, чего совершенно недостаточно, ибо, как предписывают методические указания по учету ЗМУ (1972, 1980, 1990), их должно быть «достаточно большое число».
Длина суточного хода зверя в условиях природы чрезвычайно изменчивый показатель. Например, наблюдения в течение 11 дней показывают, что длина суточного хода отдельной особи изменяется более чем в 15 раз (Козлов, 1976). Насытившись, куница прекращает охоту и залегает на отдых. В сильные морозы или метели, а также при наличии запасов корма куница по нескольку дней не выходит из убежища (Волков, 1969).
Многодневные тропления куницы в Кировской области показали, что длина суточного наследа у этих зверьков колебалась от 0 до 3,9 км. Один из самцов в течение трех дней почти не давал следа, обитая возле задавленного зайца (Плешак, 1977). Исследования лаборатории лесного охотоведения ВНИИЛМ показали, - писал П.Б. Юргенсон (1965), - что в разных угодьях даже одного приписного хозяйства приходится применять разные переходные коэффициенты, иначе неизбежны большие просчеты (например, при учете лосей).
Анализируя формулу А.Н. Формозова, на использовании которой основан метод зимних маршрутных учетов, B.C. Смирнов (1969) писал: «Разносторонняя критика формулы Формозова объясняется тем, что результаты, получаемые с ее помощью, часто оказываются неправдоподобными. Причину неверных результатов надо искать только в одном: неверные числовые величины S и d, подставляемые в формулу. Число S должно быть количеством пересечений тех самых следов, длина которых нам известна. Но измерить длину каждого пересеченного на маршрутах наследа совершенно невозможно». Избегая эту трудность в практике проведения зимних маршрутных учетов, учетчики вытрапливают лишь небольшую часть наследов и, опираясь на статистические соображения, предполагают, что все остальные наследы имеют такую же в среднем длину, как и вытропленные. Однако на этом пути появляются свои трудности, преодоление которых чрезвычайно сложно.
Чтобы определить объем выборки в случае определения длины суточного хода, нужно знать влияние многих факторов, в частности процент особей, не дававших следа вообще, то есть не выходивших из убежища, с лежки. Таковые всегда есть. Путь определения этого процента по ряду видов, а следовательно, и размах изменчивости средней длины суточного хода остается неясным, без которой определение необходимого объема выборки затруднен.
Многие исследователи считают получение средних величин длины суточных наследов, реально соответствующих действительности, невыполнимым делом. В частности, такого мнения придерживался крупнейший знаток экологии соболя В.В. Тимофеев (1973).
По своей сути длина суточного наследа есть не что иное, как показатель двигательной активности животного. Во ВНИИОЗе предприняли попытку определить показатель активности другим путем.
Другой путь определения пересчетного коэффициента. Как звестно, существуют два способа определения пересчетного коэффициента:
1. Методом тропления.
2. Сопоставлением учета зверей на пробных площадях и маршрутах, если комбинированный учет проведен в одно время и в одних местах.
Разработан метод определения пересчетных коэффициентов (Агафонов и др., 1983, 1986, 1988), в котором воедино совмещен многодневный маршрутный учёт с окладным способом определения плотности зверей на учетных площадках (рис.6). Единовременное совмещение осуществляется путем использования в качестве замкнутого маршрута периметра учетных площадок. При этом получаем возможность рассчитывать, сколько следов приходится на одного находившегося на учетной площадке зверя.
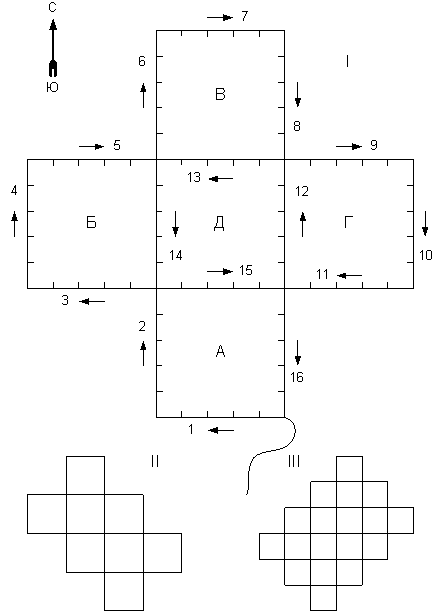
Рис. 6. Формы маршрута, дающие возможность одному человеку обойти все учётные плошадки, не проходя своим следом, и возвратиться в исходную точку.
I – «Крестообразная форма», использовавшаяся С.А. Корытиным. А, Б, В, Г, Д –учётные площадки. Извилистая линия – начальный отрезок маршрута; 1 – 16 – номера последовательно проходимых участков. Риски на маршруте – вехи.
II – Форма маршрута, предложенная В.А. Агафоновым. Даёт возможность обойти 8 учётных площадок.
III – Форма маршрута, дающая возможность обойти 18 учётных площадок
Этот способ дает возможность полнее определять запасы зверей, сравнительно прост в исполнении, менее трудоемок. При этом имеем общероссийские рекомендации по зимнему учету охотничьих животных (1990).
Ниже приводится описание деталей способа. Закладывается постоянный маршрут, идущий вокруг каждой из нескольких учетных площадок. Определяется протяженность маршрута. Для снижения трудозатрат целесообразно применить одну из форм, приведенных на рис.6, которые дают возможность огибать несколько учетных площадок, не проходя и метра старой дорогой.
Форма 1 (крестообразная) дает возможность при длине маршрута в 8 км обогнуть 5 учетных площадок площадью по 25 га (о возможности её применения упоминал B.C. Смирнов (1965). Форма 2, предложенная В.А. Агафоновым, позволяет обойти 8 таких же учетных площадок при длине маршрута 12 км. Форма 3 дает возможность вести учет на 18 учетных площадках при длине хода 24 км (фигуры этих площадок по существу являют собою комбинации «восьмерок», применявшиеся при энтомологических сборах).
| К= | 100 см | (1), |
| АХ |
где К - пересчетный коэффициент;
С - число особей на всех учетных площадках к началу учета;
М - периметр учетных площадок в километрах;
А - площадь всех учетных площадок в га;
X - среднее число следов по периметру за 5 дней наблюдений.
Формула (1) получена путем преобразования общепринятой формулы «Методические указания по организации и проведению зимнего маршрутного учета охотничьих животных в РСФСР» (1980):
К = Р:П (у) (2),
где Р - плотность зверей;
П (у) - показатель учета.
Р = (С:А) х 1000 (3); П (у) = (X 10):М (4).
Подставляя значения формул 3 и 4 в формулу 2, получаем формулу 1
Применительно к пробным площадкам и форме маршрута (рис. 6) формула (2) примет следующий вид.
| К= | 100 см | = | 100хСх10 | = | 8С | (2) |
| АХ | 125Х | Х |
С 1984 г. предложенный метод стал испытываться, при содействии В.А.Кузякина, в рамках научно-производственного эксперимента по системе Госохотучета в Ленинградской, Кировской, Костромской, Ярославской, а с 1985 г. дополнительно в Вологодской и Пермской областях (Агафонов, Корытин, Соломин, 1986). Как видно из сравнения пересчетных коэффициентов, полученных путем учета на замкнутом маршруте, и пересчетных коэффициентов по результатам тропления следов, последние значительно меньше, чем первые (табл. 9).
Пересчетные коэффициенты, полученные путем тропления, не всегда могут корректно применяться при обработке данных зимнего маршрутного учета с предварительным затиранием следов из-за реакции зверей на лыжню (Корытин, Соломин, 1982, 1988). Кроме того, существующая практика расчета средней длины суточного хода приводит к смещенной (завышенной) ее оценке, а несмещенная оценка, предложенная Н.Г. Челинцевым (1989), может оказаться неустойчивой (Соломин, 1988).
Таблица 9
Пересчетные коэффициенты (ПК), полученные на основе учета животных на замкнутом маршруте и методом тропления
| Виды | ПК, полученные путем учета на замкнутых маршрутах | ПК Госохотучета, полученные путем тропления | |
| 1984 г. | 1985 г | ||
| Заяц-беляк | 1,90(0,17) | 1,82(0,18) | 1,35 |
| Белка | 5,17(0,57) | 5,93(1,07) | 3,75 и 4,35 в 1985 г. |
| Лось | 1,97(0,40) | 1,63(0,36) | 0,72 |
| Куница | 1,91 (0,83) | 2,20(0,62) | 0,56 |
| Горностай | 3,92(0,87) | 2,40(0,57) | 1,40 |
Уточненные пересчетные коэффициенты, полученные путем учета на замкнутом маршруте, позволяют полнее определить запасы промысловых зверей и, следовательно, увеличить объем добываемой пушнины, эффективнее хозяйствовать в природе. Кроме того, способ дает выигрыш в трудозатратах: работу ведет один человек, способ прост в исполнении, позволяет стандартизировать труд.
Предложенный метод (Агафонов и др., 1983, 1986, 1988) был использован после испытаний в Методических указаниях по зимнему маршрутному учету (1990). Он оказался пригодным не только на равнинной местности, но при соблюдении определенных условий и в угодьях Западного Кавказа (Ромашин, 2007).
По приказу Министерства природных ресурсов и экологии РФ от 11 января 2012 г. № 1 несколько упрощена процедура проведения ЗМУ. Указано, что следует фиксировать количество пересечений следов данного вида копытных животных только по следующим группам категорий среды обитания: «лес», «поле», «болото». Существенным облегчением для исполнителей ЗМУ является избавление от работ по троплению суточного наследа животных. В приказе приведены готовые пересчетные коэффициенты для отдельных видов копытных животных, белки и волка с дифференцировкой для каждого вида животного по всем регионам и федеральным округам РФ, а для каждого вида пушных животных приведены усредненные коэффициенты для всей территории РФ.
Подводя итог обсуждения по следовой активности охотничьих животных, можно сделать следующие выводы.
1. Следовая активность животных в разные периоды суток видоспецифична. У изучавшихся ночных видов максимальное количество следов наблюдается в вечернее и ночное время, у белки – в дневное время.
Так, для зайца-беляка минимум следовой активности падает на 13 часов, а максимум – на 21 час. Для мышевидных грызунов характерны два пика следовой активности: первый падает на вечер, второй - на раннее утро и выражен в меньшей степени.
У лисицы изменения следовой активности менее контрастны: минимум, как у мышевидных и зайца-беляка, приходится на 13 часов, максимум - на 20 часов; ярко выражен первый пик, вечерний, и незначительно - раннеутренний.
Для лося характерна одновершинная кривая следовой активности с пиком в 18 часов.
Кабану присущ также одновершинный цикл с максимумом в 19 часов и длительным периодом спада активности, продолжающимся с полуночи до 8 часов утра.
Имеется прямая связь между следовой активностью в разные периоды суток у лисицы с зайцем-беляком и мышевидными грызунами, у последних - с лаской.
Существует прямая связь следовой активности хищника и жертвы. Видовое разнообразие следов изменяется в разные периоды суток в соответствии с их общей численностью: чем следов больше, тем большему числу видов они принадлежат.
2. Следовая активность зверей одного вида в одном и том же урочище может иметь по дням колоссальную разницу: по грызунам она в некоторых случаях достигает ста, двухсот и более раз; по хищникам и копытным максимальные различия исчисляются десятками раз, в пределах сотни. В этих пределах показатель учета может варьировать и на коротких промежутках времени, когда численность зверей данного вида не могла измениться существенно.
Динамика изменений следовой активности по дням у зверей разных видов на одном и том же постоянном маршруте иногда имеет сходство, но большей частью изменения происходят независимо друг от друга. Однако, если брать периоды по неделе и более, сходство прослеживается чаще. В изменениях следовой активности по дням у зверей какого-либо вида может возникать определенная ритмичность, но она не постоянна и сменяется аритмичными колебаниями или ритмикой с иным шагом. Для хищников характерна более равномерная следовая активность, чем для животных из разряда жертв. Численное соотношение следов хищников и жертв может меняться по годам в значительной степени.
3. Уровень следовой активности по месяцам снежного периода у изучавшихся животных весьма изменчив, колеблется по годам, однако носит печать видовой специфичности. Можно полагать, что в Кировской области для зайца-беляка (при наличии вариаций) более характерно постепенное снижение числа следов с осени вплоть до февраля, а в марте - скачок вверх; для белки - обилие следов по осени, снижение их количества зимой - в декабре, январе и возрастание к весне - в феврале и особенно в марте; для лисицы – повышение числа следов в зимние месяцы - в декабре, январе, феврале и более низкий уровень в ноябре и марте; для ласки также характерно усиление следовой активности в серединные месяцы снежного периода. У зверей, спаривание которых падает на снежный период, максимальное количество следов встречается в тот месяц, когда происходит гон. Следовая активность в разные годы и в различных районах может колебаться по каждому виду в значительной степени.
4. Следовая активность сравниваемых видов в расчете на одну особь в Подмосковье была выше до трех и более раз, чем в Кировской области, где численность зверей этих видов была ниже. Ритмика следовой активности по дням у некоторых животных, например у белки, на территориях, удаленных друг от друга (даже на расстоянии 800 км), была сходной. При сравнении подекадных изменений сходство обнаруживается для большего числа видов.
5. Имеются видовые особенности использования территории. Для лисицы характерно наличие регулярных переходов. Есть также постоянные, но иногда угасающие переходы: ими лисицы ходят не каждый год. Для горностая характерно переложное пользование территорией. Обычно ни один из четко выраженных переходов горностая в какой-либо зимний сезон на будущий год не сохранялся.
Пространственное размещение горностая менялось по годам, но за ряд лет вся территория была использована полностью.
У ласки переложное использование территории выражено в меньшей степени, чем у горностая: она более приурочена к определенным местам. Интенсивная «эксплуатация» по годам какого-либо участка местности зайцем-беляком постепенно возрастает, а затем постепенно падает, возможно, в связи с истощением кормовой базы.
Имеются места, где следы животных из года в год встречаются в большем количестве. Эти «общие переходы», «постоянные дороги» зверей не абсолютно постоянны, время от времени забрасываются, а затем снова возобновляются. Видовой состав зверей, пользующихся общим переходом, нестабилен, меняется по годам. В пределах каждой стации находится участок, наиболее предпочитаемый животными для переходов. Скопление следов по разным стациям видоспецифично.
Характер распределения следов на местности у разных зверей в зависимости от их численности меняется по-разному. Если при возрастании численности лося, ласки площадь, занятая их следами, увеличивалась, то для белки и мышевидных грызунов наблюдалось обратное явление: при высокой численности общая площадь, занятая следами этих зверьков, уменьшалась, но росла их плотность на определенных, вероятно, наиболее кормных участках. В распределении следов на местности у изучавшихся хищников и жертв имеется некоторое сходство, но оно не разительно. Между отдельными видами хищников и жертв схожесть может быть более определенной или отсутствовать совсем в зависимости от наличия экологических связей.
Картина пространственного размещения следов, а следовательно, и использования зверями территории меняется, пульсирует. Среди изучавшихся видов наиболее стабильно расположение следов в пространстве у лисицы, белки и мышевидных грызунов, более изменчиво у мелких куньих - ласки и особенно горностая, а также у зайца-беляка.
6. Реакция на лыжню, отвороты от нее присущи не только зверям, которых принято считать осторожными. Процент отворотов варьирует, но для каждого вида характерен свой количественный уровень. Соболю в глухих районах присуще усиленное мечение лыжни мочой и экскрементами. Реже метят лыжню соболи, подходившие и уходившие на прыжках, чаще - шедшие шагом. По месяцам снежного периода число отворотов варьирует, но для всех изучавшихся видов характерно возрастание их числа с осени к середине зимы (январю), а потом вплоть до марта - снижение. У хищников кривая более полога, чем у мышевидных и зайцеобразных. У тех и у других она коррелирует с температурой воздуха. Возможно, что увеличение процента отворотов вызвано ухудшением экологических условий существования, ростом стресса, уменьшение - смягчением пресса среды.
Процент отворотов коррелирует со следовой активностью в течение суток: чем больше следовая активность, тем больше процент отворотов. С возрастанием активности растет вероятность встречи с особями своего и других видов; можно полагать, что это связано с эмоциональной напряженностью, которая стимулирует осторожность, а с ней растет и процент отворотов. Играет роль рекреационный фактор: звери Вятских лесов пугаются лыжни чаще, чем в Подмосковье. Звери относятся к лыжне смелее в своих коренных, наиболее свойственных виду угодьях. Неравномерное расположение отворотов на маршруте указывает на влияние дополнительных, кроме самой лыжни, факторов.
7.Двух- и трехдневные маршрутно-окладные методы учета зверей дают заниженные результаты. Точность существенно повышается при пятидневных учетах. Численность зверей на небольших территориях (25-125 га) даже таких оседлых видов, как белка и заяц-беляк, может колебаться на протяжении 1-2 месяцев за счет откочевок и подкочевок в значительных пределах: до двух и более раз. Колебания происходят более или менее постепенно (при наличии мелких скачков), что указывает на неслучайный характер этих передвижений.
Таким образом, для получения достоверных результатов учета численности животных следует рекомендовать следующие поправки:
- Применявшийся метод четырехразового учета следов в течение суток на одном и том же маршруте с фиксацией времени прохождения отдельных его участков вполне пригоден для изучения следовой активности диких зверей в разные периоды дня и ночи, без знания которой не может быть точным авиаучет животных.
- Для сравнения относительной численности животных на удаленных друг от друга территориях зимние маршрутные учеты целесообразно производить в одни и те же дни, поскольку, как показали исследования, возможны синхронные изменения следовой активности зверей разных популяций.
- При проведении зимних маршрутных учетов интервал между затиранием следов и собственно учетом должен быть равным суточному циклу двигательной активности животных, то есть 24 часам. Нельзя проводить учеты после пороши. Это ведет к искажению результатов.
- Результаты маршрутных учетов, пересчетные коэффициенты следует представлять не только в виде средних цифр, но и с показателями изменчивости ряда, поскольку следовая активность чрезвычайно вариабельна как по отдельным дням, так и по месяцам снежного периода.
- Для нейтрализации влияния лыжни на результаты учетных работ предлагаем введение учетной полосы, перекрывающей поле эмоционального воздействия лыжни на животное. Полоса имеет ширину 5 метров, а лыжня проходит по её середине.
- Применяемый метод ежедневных учетов на одном и том же маршруте с большим количеством постоянных ориентиров может быть использован для изучения ритмики следовой активности и факторов, влияющих на нее, для исследования пространственного распределения животных, зависимости его от стаций, численности населения вида и других причин, позволяет фиксировать расположение следов зверей с точностью до одного сантиметра.
Анализ огромного массива материалов по учетам и оценке ресурсов диких животных в стране позволяет заключить, что наиболее целесообразна следующая структура мониторинга охотничьих животных:
1. Ежегодный, двукратный опрос охотников–корреспондентов по широкому спектру видов и вопросов по системе Службы «урожая» ВНИИОЗ.
2. Проведение (ЗМУ) по системе Госохотучета по видам, подлежащим учетам в зимних условиях, один раз в два-три года.
3. Проведение специализированных областных видовых учетов, не проходящих по системе ЗМУ (горные, полуводные, зимоспящие, «летние» виды и т.д.), в промежутке между годами ведения ЗМУ (по каждому виду или группе видов один раз в 3-5 лет).
4. Использовать данные Летописи природы системы заповедников для получения базовых данных по численности видов, отнесенных к объектам охоты, а также по редким и исчезающим видам животных.
Такая система взаимодействия трех источников (службы Госохотучета, Службы «урожая», Летописи природы) обеспечит взаимоконтроль, повышение точности оценок, снижение трудовых, материальных и денежных затрат на ведение мониторинга охотничьих видов животных.
Объектами мониторинга охотничьих животных являются: лось, кабан, косуля, бурый медведь, соболь, лесная куница, хорьки, колонок, норки, выдра, горностай, волк, лисица, корсак, рысь, белка, сурок, бобр, ондатра, зайцы, глухарь, тетерев, рябчик, водоплавающие птицы и другие виды.
Характеристика объекта. В пределах региона эти виды встречаются не повсеместно, но наиболее многочисленны в типичных для каждого из них местах обитания с хорошей кормовой базой, благоприятными гнездопригодными условиями и низкой (умеренной) интенсивностью охоты. Состав видов, их соотношение и обилие особей в территориальных группировках в различные сезоны года и разные годы подвержены изменчивости.
Изменение структуры видового состава охотничьих животных по сравнению с фоновым состоянием свидетельствует об изменениях в среде их обитания. Плотность населения животных является отражением экологической емкости ценоза. Снижение численности большинства видов млекопитающих может быть результатом ухудшения кормности угодий и условий для размножения, чрезмерного преследования и пр. Стабилизация и рост численности животных свидетельствует о возможности их сопротивления действию внешних факторов и самовосстановлению после снятия этого воздействия и других причин. Ответная реакция на воздействие может быть зафиксирована не по каждому виду в течение одного года.
Некоторые виды охотничьих животных могут использоваться в качестве тест-объектов для оценки загрязнения среды, степени ее трансформации в городах и техногенных зонах.
Оборудование и материалы. Требуются физические и административные карты региона, района и охотхозяйства; планшет учетчика; карточки учета; компас; лыжи охотничьи; 2-местная лодка надувная; стандартные опросные анкеты (по 2 в год на каждого респондента); репрезентативная (10-15 человек на административный район) корреспондентская сеть; конверты (по 4 в год на каждого респондента); писчая бумага; ручки; карандаши; компьютер с базой данных, цветные и черно-белые картриджи; CD-диски, флэшкарты.
Методика исследований. Как основную на учетных полигонах для учета охотничьих животных применяют методику многодневного оклада на замкнутых маршрутах (Агафонов и др., 1983; «Методические указания по организации, проведению и обработке данных ЗМУ охотничьих животных России» (2008) и различные повидовые методы). Постоянная пробная площадка (ППП) размером 800 га с квадратами 1000х1000 м предназначена для учета копытных, средних и крупных хищников (рис.7).
Работы проводят два учетчика. Для учетов белки, зайцев, горностая, колонка, хорьков используется ППП размером 200 га с квадратами 500х500 м. Учеты проводят 1-2 учетчика. Работы ведутся 4 дня подряд. В первый день затирают все старые следы. В каждый из последующих трех дней проходят по маршруту, затирают и одновременно отмечают каждый день на новой схеме своего маршрута все вновь появившиеся пересечения следов с регистрацией их направления и вновь появившиеся подходы зверей к лыжне. Время учета – конец января-февраль, до начала гона хищных, зайцеобразных и грызунов.
Околоводных животных (бобр, ондатра, выдра, норка) учитывают специальными методами. Сущность маршрутного учета бобра заключается в выявлении и подсчете в водоеме (река или замкнутый водоем) всех бобровых поселений с глазомерной оценкой количества и характера распределения следов деятельности зверей (погрызы, жилища, плотины, вылазы и пр.) для оценки мощности поселения (слабое поселение - 1-2 зверя, среднее – 4, в том числе сеголетки, сильное – 5 и более зверей).
Дата учета "_____"___________20____г.
Погода в день учета: температура от_______ до _______, осадки________
_________________ветер (сила, направление)_________________________
Высота снежного покрова ________ см. Характер снега (рыхлый, плотный, с коркой, наст и т.п.)______________________________________________
СХЕМА ОКЛАДА
| Условные знаки для оклада в 800 га Волк Во Кабан Ка Косуля Кс Куница Ку Лисица Ли Лось Ло Олень благор Об (изюбрь Из Марал) Росомаха Ро Рысь Ры Соболь Со® | ||||||||
| Условные знаки для оклада в 200 га Белка Бе Горностай Го Заяц-беляк Зб Заяц-русак Зр Колонок Ко Хорек Хо | ||||||||
| Примеры пометок | ||||||||
| Прошли 3 лося |  | Порядок обхода площадок двумя учетчиками | ||||||
| Подход волка |  | 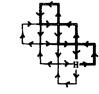 |  | - движение 1-го учетчика | ||||
 | - движение 2-го учетчика | |||||||
| Выход лисицы на лыжню |  | Н | - начало хода | |||||
Рис. 7. Схема учета охотничьих животных на замкнутых маршрутах
Признаки молодняка: отпечатки ступней задних лап длиной не более 12-13 см, ширина следов резцов на погрызах веток не более 5-6 мм. Годовики, соответственно имеют 13,5-16 см и 6,5-7,8 мм.
Установление границ поселений (в одном поселении может быть несколько плотин и жилищ). Между соседними поселениями должно быть не менее 200 м, лишенных свежих следов жизнедеятельности. Учет проводят осенью до ледостава.
Ондатру в замкнутых водоемах (озера, пруды, болота) учитывают по семейным хаткам на площадках размером 150-200 га. При обитании ондатры в норах её учитывают по этим убежищам на маршрутах протяженностью до 10 км, заложенных по берегам водоемов. При этом границы поселений норной ондатры устанавливают по следам жизнедеятельности (вылазы, кормовые столики, погрызы, уборные). Учет проводят в течение месяца до ледостава.
Сурка учитывают визуально на маршрутах (семьи) и на площадках (особей в семьях) в период массового выхода сеголетков (через 10-15 дней после первого их появления на поверхности) (Машкин, Челинцев, 1989).
Норку и выдру учитывают по следам зверей (отпечатки лап, экскременты, вылазы и пр.) на маршрутах вдоль береговой линии (лучше по снегу). Расстояние в 250 м без набродов норки и других следов жизнедеятельности считается границей между индивидуальными участками зверьков. Индивидуальныйcуточный ход норки в период замерзания водоемов достигает 1-3 км, у выдры – 3-8 км. Учет проводят поздней осенью по первому снегу, можно и по льду до образования пустоледиц.
Учет медведя проводят по индивидуальным отпечаткам (измеренным мерной линейкой) плантарной мозоли передней лапы зверя на маршрутах (лесные дороги, просеки, противопожарные полосы, окраины овсяных полей, берега рек и т.д.). При отсутствии таковых в местах пребывания медведей целесообразно соорудить контрольно-следовые полосы. Протяженность маршрута до 15 км. Учет проводят осенью до наступления заморозков.
Охотничьих птиц учитывают по общепринятой методике («Методические указания по проведению осеннего маршрутного учета боровой и полевой дичи». М.,1989). Учетные маршруты закладывают равномерно по всей территории с пропорциональным охватом всех угодий и прохождением по азимуту, узким квартальным просекам или дорогам. Нельзя закладывать маршруты по широким (более 4 м) дорогам и просекам, окраинам полей, опушкам, вдоль водоемов.
Учет на учетной ленте длиной 10-12 км проводит один учетчик в период с августа по октябрь включительно. В полевом журнале (табл. 10) указываются данные по каждой встрече.
Водоплавающих птиц на замкнутых водоемах подсчитывают визуально (всех увиденных). На линейных водоемах визуальный учет осуществляется на маршрутах в 10-12 км и по ширине обследованного участка реки, старицы.
Проводится полный учет всех встреченных взрослых птиц и выводков (по видам) на модельных водоемах полигона.
Таблица 10
Ведомость летне-осеннего учета куриных птиц
Район_______________ Полигон_____________________________
Общая протяженность маршрутов, км ________________________;
в том числе лес________км, поле_________км, болото________км
| Дата | Вид | Количество птиц | Расстояние вспугивания, м | Характер угодий | Примечание* |
*- в примечании указывают пол, возраст птиц (в случае определения), поведение птиц и т.д.
Сведения о встреченных птицах заносятся в полевой журнал (табл.11). Время учета - июль-первая половина августа.
Таблица 11
Ведомость летнего учета водоплавающих птиц
Район___________________ Полигон________________________
Период учета_____________________________________________
| Название водоема | Площадь, га | Учтено птиц | Примечание | |||
| вид | всего | в том числе | ||||
| взрослых | молодых | |||||
Опросный метод. Ежегодно два раза в год респондентам высылаются специализированные для каждой природно-климатической зоны анкеты (сведения А-1 для осени-зимы и А-2 для весны-лета) для относительной оценки (много, средне, мало) обилия охотничьих животных, урожайности их основных кормов, размножения различных видов и сравнительной с предыдущим сезоном оценки (больше, меньше, не изменилось) (см. приложение).








