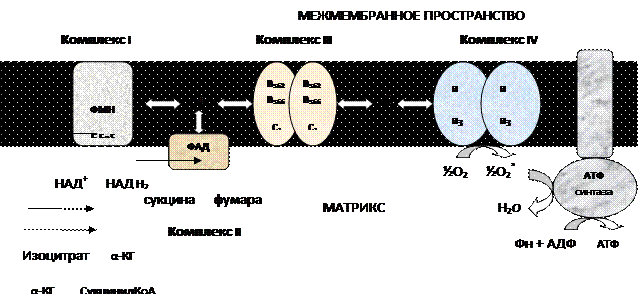
Дыхательная цепь окислительного фосфорилирования состоит из 4 белковых комплексов, встроенных во внутреннюю мембрану митохондрий и небольших подвижных молекул убихинона и цитохрома С, которые циркулируют в липидном слое мембраны между белковыми комплексами.
Комплекс I – НАДН2 дегидрогеназный комплекс – самый большой из дыхательных ферментных комплексов –в качестве коферментов содержит ФМН и 5 железо-серных (Fe
2S
2 и Fe
4S
4) белков.
Комплекс II – СДГ. В качестве коферментов содержит ФАД и железо-серный белок.
Комплекс III – Комплекс b-c1 (фермент QH2 ДГ), существует в виде димера. Каждый мономер содержит 3 гема, связанных с цитохромами b
562, b
566, с
1, и железо-серный белок.
Комплекс IV – Цитохромоксидазный комплекс существует в виде димера. Каждый мономер содержит 2 цитохрома (а и а
3) и 2 атома меди.
Коэнзим Q (убихинон). Липид, радикал которого у млекопитающих образован 10 изопреноидными единицами (Q
10). Убихинон переносит по 2Н
+ и 2е
-. убихинон ↔ семихинон ↔ гидрохинон
Цитохром с. Периферический водорастворимый мембранный белок с молекулой гема.
Хемиосмотическая теория Митчелла Для объяснения механизма окислительного фосфорилирования в 1961 году Митчеллом была предложена хемиосмотическая теория, которая включала четыре независимых постулата, касавшиеся
функции митохондрий:
1.Внутренняя мембрана митохондрий непроницаема для всех ионов.
2.Она содержит ряд белков-переносчиков, осуществляющих транспорт необходимых метаболитов и неорганических ионов.
3. При прохождении электронов по дыхательной цепи внутренней мембраны происходит перемещение Н+ из матрикса в межмембранное пространство.
4. При достаточно большом протонном градиенте протоны начинают «течь» через АТФ-синтетазу, что сопровождается синтезом АТФ.
 2015-10-22
2015-10-22 1954
1954







