 2018-01-21
2018-01-21 4510
4510Мембранный потенциал (МП) представляет собой разность потенциалов между наружной и внутренней поверхностями мембраны возбудимой клетки в условиях ее покоя. В среднем у клеток возбудимых тканей МП достигает 50 – 80 мВ, со знаком минус внутри клетки. Исследование природы мембранного потенциала показало, что во всех возбудимых клетках (нейроны, мышечные волокна, миокардиоциты, гладкомышечные клетки) его наличие обусловлено преимущественно ионами К+. Как известно, в возбудимых клетках за счет работы Na-K-насоса концентрация ионов К+ в цитоплазме в условиях покоя поддерживается на уровне 150 мМ, в то время как во внеклеточной среде концентрация этого иона обычно не превышает 4 – 5 мМ. Это означает, что внутриклеточная концентрация ионов К+ в 30 – 37 раз выше, чем внеклеточная. Поэтому по градиенту концентрации ионы К+ стремятся выйти из клетки во внеклеточную среду. В условиях покоя, действительно, существует выходящий из клетки поток ионов К+, при этом диффузия осуществляется по калиевым каналам, большая часть которых открыта. В результате того, что мембрана возбудимых клеток непроницаема для внутриклеточных анионов (глутамата, аспартата, органических фосфатов), на внутренней поверхности мембраны клетки вследствие выхода ионов К+ образуется избыток отрицательно заряженных частиц, а на наружной – избыток положительно заряженных частиц. Возникает разность потенциалов, т. е. мембранный потенциал, который препятствует чрезмерному выходу ионов К+ из клетки. При некотором значении МП наступает равновесие между выходом ионов К+ по концентрационному градиенту и входом (возвратом) этих ионов по возникшему электрическому градиенту. Мембранный потенциал, при котором достигается это равновесие, получил название равновесного потенциала. Помимо ионов К+ определенный вклад в создание мембранного потенциала вносят ионы Na+ и Сl. В частности, известно, что концентрация ионов Na+ во внеклеточной среде в 10 раз больше, чем внутри клетки (140 мМ против 14 мМ). Поэтому ионы Na+ в условиях покоя стремятся войти в клетку. Однако основная часть натриевых каналов в условиях покоя закрыта (относительная проницаемость для ионов Na+, судя по экспериментальным данным, полученным на гигантском аксоне кальмара, в 25 раз ниже, чем для ионов К+). Поэтому в клетку входит лишь небольшой поток ионов Na+. Но и этого достаточно, чтобы хотя бы частично компенсировать избыток анионов внутри клетки. Концентрация ионов Сl- во внеклеточной среде также выше, чем внутри клетки (125 мМ против 9 мМ), и поэтому эти анионы также стремятся войти в клетку, очевидно, по хлорным каналам.
Мембранный потенциал
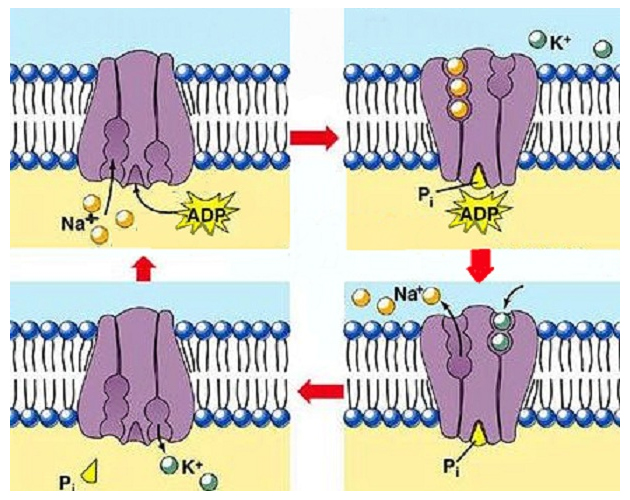
Мембранный потенциал покоя крупных нервных волокон, когда по ним не проводятся нервные сигналы, составляет около -90 мВ. Это значит, что потенциал внутри волокна на 90 мВ отрицательнее, чем потенциал внеклеточной жидкости снаружи волокна. Далее мы объясним все факторы, определяющие уровень этого потенциала покоя, но прежде необходимо описать транспортные свойства мембраны нервного волокна для ионов натрия и калия в условиях покоя. Активный транспорт ионов натрия и калия через мембрану. Натрий-калиевый насос. Вспомним, что все клеточные мембраны организма имеют мощный Na+/K+-Hacoc, постоянно выкачивающий ионы натрия наружу клетки и закачивающий внутрь нее ионы калия. Это электрогенный насос, поскольку положительных зарядов наружу перекачивается больше, чем внутрь (3 иона натрия на каждые 2 иона калия, соответственно). В результате внутри клетки создается общий дефицит положительных ионов, ведущий к отрицательному потенциалу с внутренней стороны клеточной мембраны. Na+/K+-Hacoc создает также большой градиент концентрации для натрия и калия через мембрану нервного волокна в покое: Na+ (снаружи): 142 мэкв/л Na+ (внутри): 14 мэкв/л К+ (снаружи): 4 мэкв/л К+ (внутри): 140 мэкв/л Соответственно, отношение концентраций двух ионов внутри и снаружи составляет: Na внутри / Na снаружи — 0,1 К внутри / -К снаружи = 35,0
Утечка калия и натрия через мембрану нервного волокна. На рисунке показан канальный белок в мембране нервного волокна, называемый каналом калий-натриевой утечки, через который могут проходить ионы калия и натрия. Особенно существенна утечка калия, поскольку каналы более проницаемы для ионов калия, чем натрия (в норме примерно в 100 раз). Как обсуждается далее, это различие в проницаемости чрезвычайно важно для определения уровня нормального мембранного потенциала покоя.
Таким образом, основными ионами, определяющими величину МП, являются ионы К+, покидающие клетку. Ионы Na+, входящие в клетку в небольших количествах, частично уменьшают величину МП, а ионы Сl-, также входящие в клетку в условиях покоя, в определенной степени компенсируют это влияние ионов Na+. Кстати, в многочисленных экспериментах с различными возбудимыми клетками установлено, что чем выше проницаемость клеточной мембраны для ионов Na+ в условиях покоя, тем ниже величина МП. Для того чтобы МП поддерживался на постоянном уровне, необходимо поддержание ионной асимметрии. Для этого, в частности, служат ионные насосы (Na-K-насос, а также, вероятно, Сl-насос) которые восстанавливают ионную асимметрию, особенно после акта возбуждения. Так как этот вид транспорта ионов активный, т. е. требующий затраты энергии, то для поддержания мембранного потенциала клетки необходимо постоянное наличие АТФ.
НА мембранный потенциал нейронов, а также многих других клеток, влияют изменения внеклеточной концентрации калия, но не хлора. Обратимся к модели идеальной клетки. Сделаем допущение (для рассмотрения данной ситуации), что объем внеклеточной жидкости бесконечно велик, и что перемещение ионов не влечет за собой значительного изменения концентрации ионов вне клетки. На рис.1.2А показаны изменения внутриклеточного ионного состава и мембранного потенциала, вызванные повышением внеклеточного уровня калия с 3 до 6 ммоль. Для сохранения первоначальной осмолярности, одновременно с добавлением 3 ммоль калия из раствора удалили 3 ммоль натрия, в результате чего общая концентрация растворенных ионов осталась на уровне 240 ммоль. Увеличение внеклеточной концентрации калия приводит к снижению его трансмембранного градиента, движущего ионы из клетки наружу. При этом на начальном этапе мембранный потенциал не меняется. В результате происходит перенос суммарного положительного заряда внутрь клетки. Вследствие накопления положительного заряда на внутренней поверхности мембраны она деполяризуется. Ионы натрия благодаря деполяризации выходят из состояния равновесия и начинают двигаться внутрь клетки. Перемещение ионов калия и хлора продолжается до тех пор, пока не будет достигнуто новое равновесное состояние, основанное на новом соотношении концентраций и новом уровне мембранного потенциала, в данном примере - 68 мВ.
Вход калия и хлора сопровождается током в клетку некоторого количества воды, приводящим к небольшому увеличению объема клетки. В процессе достижения нового состояния равновесия концентрация калия возрастает с 90 до 91 ммоль, хлора - с 4 до 7,9 ммоль, а объем клетки увеличивается на 3,5%. На первый взгляд кажется, что количество вошедшего в клетку хлора превышает количество калия, но представим себе, каковы были бы эти концентрации, если бы объем клетки не увеличился: концентрации обоих ионов были бы выше на 3,5%. Следовательно, вместо 7,9 ммоль концентрация хлора была бы 8,2 ммоль, а концентрация калия была бы на уровне 94,2 ммоль, то есть увеличение обеих концентраций составило бы 4,2%. Таким образом, сначала калий и хлор входят в клетку в приблизительно равных количествах (за исключением незначительного количества, необходимого для изменения заряда на мембране), а уже затем вода втекает в клетку, снижая концентрации обоих ионов до окончательного уровня.
4. Лейкоциты, их количество, функции, образование. Лейкоцитарная формула.
Лейкоцит (от др.-греч. λευκός — белый и κύτος — вместилище, тело) — белые кровяные клетки; неоднородная группа различных по внешнему виду и функциям клеток крови человека или животных, выделенная по признакам наличия ядра и отсутствия самостоятельной окраски.Образование лейкоцитов (лейкопоэз) проходит в костном мозге и лимфоузлах.
Главная сфера действия лейкоцитов — защита. Они играют главную роль в специфической и не специфической защите организма от внешних и внутренних патогенных агентов, а также в реализации типичных патологических процессов.
Все виды лейкоцитов способны к активному движению и могут переходить через стенку капилляров и проникать в межклеточное пространство, где они поглощают и переваривают чужеродные частицы. Этот процесс называется фагоцитоз, а клетки, его осуществляющие, — фагоциты.
Если чужеродных тел проникло в организм очень много, то фагоциты, поглощая их, сильно увеличиваются в размерах и в конце концов разрушаются. При этом освобождаются вещества, вызывающие местную воспалительную реакцию, которая сопровождается отеком, повышением температуры и покраснением пораженного участка.
Вещества, вызывающие реакцию воспаления, привлекают новые лейкоциты к месту внедрения чужеродных тел. Уничтожая чужеродные тела и поврежденные клетки, лейкоциты гибнут в больших количествах. Гной, который образуется в тканях при воспалении, — это скопление погибших лейкоцитов.
Виды лейкоцитов[править | править вики-текст]
Лейкоциты — собирательное понятие, введённое в XIX веке и сохраняемое для простоты противопоставления «белая кровь—красная кровь». По современным данным лейкоциты различаются по происхождению, функциям и внешнему виду. Часть лейкоцитов способны захватывать и переваривать чужеродные микроорганизмы (фагоцитоз), а другие могут вырабатывать антитела. Вследствие этого существует несколько видов деления лейкоцитов, простейший из которых основан на наличии/отсутствии специфических гранул в их цитоплазме.
По морфологическим признакам лейкоциты, окрашенные по Романовскому—Гимзе, со времён Эрлиха традиционно делят на две группы:
· зернистые лейкоциты, или гранулоциты — клетки, имеющие крупные сегментированные ядра и обнаруживающие специфическую зернистость цитоплазмы; в зависимости от способности воспринимать красители они подразделяются на нейтрофильные, эозинофильные и базофильные;
· не зернистые лейкоциты, или агранулоциты — клетки, не имеющие специфической зернистости и содержащие простое несегментированное ядро, к ним относятся лимфоциты и моноциты.
Соотношение разных видов белых клеток, выраженное в процентах, называется лейкоцитарной формулой.
Исследование количества и соотношения лейкоцитов является важным этапом в диагностике заболеваний.
Эозинофилы — лейкоциты, содержащие двудольчатое ядро и гранулы, которые окрашиваются эозином в красный цвет. Они регулируют аллергические реакции, их количество возрастает при аллергиях, а также в случаях заражения паразитическими червями (гельминтами).
Количество лейкоцитов[править | править вики-текст]
В крови взрослого человека лейкоцитов содержится в 1000 раз меньше, чем эритроцитов, и в среднем их количество составляет 4—9·109/л. У новорождённых детей, особенно в первые дни жизни, количество лейкоцитов может сильно варьировать от 9 до 30·109/л. У детей в возрасте 1—3 года количество лейкоцитов в крови колеблется в пределах 6,0—17,0·109/л, а в 6—10 лет в пределах 6,0-11,0·109/л[1][2].
Содержание лейкоцитов в крови не является постоянным, а динамически изменяется в зависимости от времени суток и функционального состояния организма. Так, количество лейкоцитов обычно несколько повышается к вечеру, после приёма пищи, а также после физического и эмоционального напряжения.
Увеличение общего абсолютного количества лейкоцитов в единице объёма выше верхней границы нормы называется абсолютным лейкоцитозом, а уменьшение её ниже нижней границы — абсолютная лейкопения.
Лейкоцитарная формула включает в себя определение относительного количества (%) нейтрофилов, лимфоцитов, эозинофилов, базофилов, моноцитов.
Исследование лейкоцитарной формулы имеет большое значение в диагностике гематологических, инфекционных, воспалительных заболеваний, а также оценке тяжести состояния и эффективности проводимой терапии. В то же время, изменения лейкоцитарной формулы не являются специфичными - они могут иметь сходный характер при разных заболеваниях или, напротив, могут встречаться непохожие изменения при одной и той же патологии у разных больных.
Лейкоцитарная формула имеет возрастные особенности, поэтому ее сдвиги должны оцениваться с позиции возрастной нормы (это особенно важно при обследовании детей).
Метод определения: микроскопия мазка крови врачом-лаборантом с подсчетом лейкоцитарной формулы на 100 клеток.
Лейкоциты (белые кровяные тельца, whitebloodcells)
По морфологическим признакам (вид ядра, наличие и характер цитоплазматических включений) выделяют 5 основных видов лейкоцитов - нейтрофилы, лимфоциты, моноциты, эозинофилы и базофилы. Кроме того, лейкоциты различаются по степени зрелости. Большая часть клеток-предшественников зрелых форм лейкоцитов (юные, миелоциты, промиелоциты, бластные формы клеток), а также плазматические клетки, молодые ядерные клетки эритроидного ряда и др. в периферической крови появляются только в случае патологии.
Различные виды лейкоцитов выполняют разные функции, поэтому определение соотношения разных видов лейкоцитов, содержания молодых форм, выявление патологических клеточных форм, описание характерных изменений морфологии клеток, отражающих изменение их функциональной активности, несет ценную диагностическую информацию.
Некоторые варианты изменения (сдвига) лейкоцитарной формулы:
Сдвиг влево (в крови присутствует увеличенное количество палочкоядерных нейтрофилов, возможно появление метамиелоцитов (юных), миелоцитов) может указывать на:
- Острые инфекционные заболевания;
- Физическое перенапряжение;
- Ацидоз и коматозные состояния.
Сдвиг вправо (в крови появляются гиперсегментированные гранулоциты) может указывать на:
- Мегалобластную анемию;
- Болезни почек и печени;
- Состояния после переливания крови.
Значительное омоложение клеток:
- Так называемый «бластный криз» - наличие только областных клеток: острые лейкозы, метастазы злокачественных новообразований, обострение хронических лейкозов;
- «провал» лейкоцитарной формулы – бластные клетки, промиелоциты и зрелые клетки, промежуточных форм нет: характерно для дебюта острого лейкоза.
5. Структура и функции нервных волокон, классификация нервных волокон. Механизм проведения возбуждения безмякотным и мякотным нервным волокнам. Законы проведения возбуждения по нервным волокнам: изолированное и двустороннее проведение, практическаянеутомляемость.
Не́рвныеволо́кна — длинные отростки нейронов, покрытые глиальными оболочками.
В различных отделах нервной системы оболочки нервных волокон значительно отличаются по своему строению, что лежит в основе деления всех волокон на миелиновые и безмиелиновые. Те и другие состоят из отростка нервной клетки, лежащего в центре волокна, и поэтому называемого осевым цилиндром (аксоном), и окружающей его миелиновой оболочкой. В зависимости от интенсивности функциональной нагрузки нейроны формируют тот или иной тип волокна. Для соматического отдела нервной системы, иннервирующей скелетную мускулатуру, обладающую высокой степенью функциональной нагрузки, характерен миелиновый (мякотный) тип нервных волокон, а для вегетативного отдела, иннервирующего внутренние органы — безмиелиновый (безмякотный) тип.

