 2014-02-03
2014-02-03 2697
2697Благодаря системам активного транспорта для 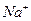 и
и 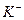 , поддерживаются стабильные и высокие градиенты концентрации этих ионов на плазмолемме любой клетки. Разница в их молекулярной концентрации между цитозолем и межклеточной средой достигается 10-20 раз. При этом, больше в цитозоли, - в межклеточной среде. При действии концентрационных градиентов входят в нее. Пассивный транспорт привел бы к ликвидации ионного неравновесия, но этому препятствует работа
, поддерживаются стабильные и высокие градиенты концентрации этих ионов на плазмолемме любой клетки. Разница в их молекулярной концентрации между цитозолем и межклеточной средой достигается 10-20 раз. При этом, больше в цитозоли, - в межклеточной среде. При действии концентрационных градиентов входят в нее. Пассивный транспорт привел бы к ликвидации ионного неравновесия, но этому препятствует работа  насоса. Он откачивает из цитозоля в межклеточную среду и закачивает в клетку. насос - энергоемкая система.
насоса. Он откачивает из цитозоля в межклеточную среду и закачивает в клетку. насос - энергоемкая система.
Компонент помпы является АТФ (источник энергии) АТФазы, которые одновременно служит и переносчиком и сопрягающим фактором. Молекулярный механизм использования энергии АТФ для работы помпы еще полностью не понят, однако, предполагается, что на внутренней стороне БМ находятся молекулярные комплексы, способные фосфорелировать за счет присоединения фосфатной группы АТФ. Фосфатная группа отщепляется от АТФ при гидролизе. Фосфорелирующий транспортный комплекс переносит, связанный с ним на наружную сторону клеточной мембраны, где обменивает на . комплекс транспортирует его внутрь клетки, где он фосфолерируется, то есть, отщепляется фосфатная группа, так как вся свободная энергия истрачена. Для следующего транспортного цикла комплексу необходимо новое фосфорелирование за счет гидролиза АТФ.
В настоящее время есть несколько гипотез, конкретизирующих участие в работе комплекса -АТФазы. Однако, ни одна из них не является общепринятой, но, все модели активного транспорта и 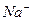 предусматривает конфирмацию перестройки -АТФазы. Эти перестройки осуществляются в пространстве БМ.
предусматривает конфирмацию перестройки -АТФазы. Эти перестройки осуществляются в пространстве БМ.
Некоторые условно полагают, что, будто помпа действует по принципу перистальтического насоса, который переменно открывает и закрывает и, расположенные в БМ по соседству с АТФазой.
Ее конформация перестройки обуславливается чередованием фосфорелирования и дефосфорелирования, обеспечивают изменение проницаемости ионов каналов, при этом, эти насосы работают в противофазе, то есть, когда канал открыт, - закрыт, и наоборот. Согласно этой гипотезе, АТФаз служит не как переносчик, а своеобразное планирование ионных каналов.
Другая гипотетическая схема действия помпы предполагает, что транспорт АТФаз работает, как переносчик. Молекула этого фермента пронизывает плазмолемму насквозь, контактируя одним полюсом с цитоплазмой, другим - с межклеточной средой. В при мембранных участках цитоплазмы находятся митохондрии, из которых выходят молекулы АТФ. АТФ выступает в контакте с БМ, следовательно, и с АТФазой. Содержание ионов Na в цитоплазме поддерживается на строго определенном уровне. АТФаза активируется ионами и , но проявляется при этом ярко выраженная векторность. Ионы действуют на нее только со стороны цитоплазмы, а - только из межклеточной среды. Вместе с тем, ее специфичность по отношению к выше, чем к , так, для включения в действие АТФазы, незаменимы, в то время, как можно заменить одновалентным катионом, причем, с рубидием транспортная система функционирует лучше, чем с . По-видимому, для ферм. характерны различные механизмы распознавания на цитоплазме стороне БМ и других катионов на ее наружной поверхности.
Экспериментально установлено, что за счет гидролиза одной молекулы АТФ осуществляется активный транспорт трех ионов и двух ионов , то есть, их сопряженному трансмембранному переносу свойственна стехиометрия (3/2). Она сохраняется вне зависимости от величины и направления концентрационного и электрического градиентов. Стехиометрия обусловливает электрогенные свойства помпы. Перенос двух внутрь клетки и трех ионов Na из нее, создает разность потенциалов, относительно межклеточной среды. При этом цитозоль имеет отрицательный заряд относительно межклеточной среды (внутри клетки анионов больше, чем катионов).
насос угнетается различными агентами, из которых наиболее активны следующие:
2. Избыток ионов 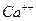 внутри клетки.
внутри клетки.
3. Дыхательные яды.
Дыхательные яды блокируют окислительное фосфорилирование в митохондриях и, соответственно, они останавливают работу любой системы активного транспорта, то есть, в их действии нет специфичности, так как они лишают свободной энергии активного транспорта. Специфичным плантатором насоса служит строфангин Г (уабаин), который является ингибитором АТФазы. Даже в концентрации, равной 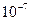 м/а, уабаин подавляет активность помпы на 50 %. Под действием гликозидов подавляется выведение из клеток, и приток в них, тогда как пассивный выход из цитозоли не изменяется.
м/а, уабаин подавляет активность помпы на 50 %. Под действием гликозидов подавляется выведение из клеток, и приток в них, тогда как пассивный выход из цитозоли не изменяется.
КАЛЬЦИЕВЫЙ НАСОС (КН)
КН обеспечивает стабильно низкий уровень 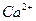 в цитозолях, в отличие от насоса, он выводит избыток ионов , не в межклеточную среду, а в органеллы (главным образом, в эндоплазматическую сеть). Поэтому, основные места локализации насоса находятся в большинстве типов клеток на внутренней мембране, а не на плазмолемме (что имело место для насоса). Достаточно детально изучена работа помпы в мембранах саркоплазматической сети, где ее активность наиболее высока.
в цитозолях, в отличие от насоса, он выводит избыток ионов , не в межклеточную среду, а в органеллы (главным образом, в эндоплазматическую сеть). Поэтому, основные места локализации насоса находятся в большинстве типов клеток на внутренней мембране, а не на плазмолемме (что имело место для насоса). Достаточно детально изучена работа помпы в мембранах саркоплазматической сети, где ее активность наиболее высока.
Источником энергии для системы активного транспорта ионов служит АТФ. Вторым и третьим компонентом работы насоса является активированная АТФаза. Кроме того, для работы АТФазы необходимо присутствие ионов 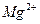 . В саркоплазматической цепи на долю АТФазы приходится около 60 % общего мембранного белка. Предполагается, что в мембране саркоплазматической сети нет другого белка, кроме АТФазы. Остальные 40 % белков принадлежат периферическим протеинам. На перенос двух ионов затрачивается одна молекула АТФ, но, при очень высоких физико-химических градиентах, это отношение изменяется с двух к одному, до одного к одному.
. В саркоплазматической цепи на долю АТФазы приходится около 60 % общего мембранного белка. Предполагается, что в мембране саркоплазматической сети нет другого белка, кроме АТФазы. Остальные 40 % белков принадлежат периферическим протеинам. На перенос двух ионов затрачивается одна молекула АТФ, но, при очень высоких физико-химических градиентах, это отношение изменяется с двух к одному, до одного к одному.
В механизме действия насоса выделяется три этапа:
Во-первых, сигналом к активному транспорту служит увеличение допустимого уровня ионов в цитозоле. Установлено, что постоянная связывания ионов кальция- АТФазой, имеет порядок 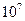 л/моль, то есть, адсорбция ионов
л/моль, то есть, адсорбция ионов 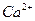 на транспортном ферменте происходит уже при концентрации ионов в цитозоли примерно моль/л. - АТФаза связывает не только ионы, но и ионы . Центры захвата ионов и АТФ локализованы на поверхности фермента. Эта поверхность обращена к цитозоле.
на транспортном ферменте происходит уже при концентрации ионов в цитозоли примерно моль/л. - АТФаза связывает не только ионы, но и ионы . Центры захвата ионов и АТФ локализованы на поверхности фермента. Эта поверхность обращена к цитозоле.
Активированная ионами транспортная АТФаза катализирует гидролиз АТФ, что является основным событием второго этапа в работе помпы. При гидролизе АТФ, от нее отщепляется концевая фосфатная группа и присоединяется к - АТФазе, которая приобретает при этом дополнительную свободную энергию равную примерно 37,4 Дж/моль. За счет этой энергии образуется фермент-фосфатный комплекс.
Третий этап работы насоса - переход ионов на противоположную сторону внутри БМ, что связано с изменением конформации АТФазы. Изменение конформации приводит к перемещению молекулы фермента в пространстве БМ. Предполагается, что происходит сдвиг подвижных групп АТФазы, на которой адсорбированы ионы. На противоположной стороне БМ транспортная АТФаза освобождается от ионов, так как ферментно-фосфатный комплекс гидрализуется, после того, как затрачивает полученную энергию на активный транспорт ионов. Вслед за гидролизом ферментно-фосфатного комплекса, происходит дефосфорилирования фермента. Возвращение - связывающих центров в исходное состояние является следствием восстановительной конформации молекулы АТФазы. Это состояние переходит в то, которое ей является свойственным в нефосфорелированном состоянии.
насос, в отличие от насоса, не проявляет электрогенных свойств, то есть, активный транспорт ионов не сопровождается образованием разности потенциалов на БМ. Неэлектрогенность помпы обусловлена высокой проницаемостью этой мембраны для многих ионов. В этой связи, мембранный потенциал, который создается переносом ионов, сразу же падает из-за утечки других ионов.

