 2018-01-21
2018-01-21 1487
1487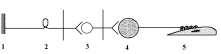 (СНС)
(СНС)
1 – рецептор; 2 – афферентный нейрон; 3 – центральный (преганглионарный) нейрон; 4 – ганглионарный нейрон (симпатический ганглий); 5 – эффектор (гладкая мышца).
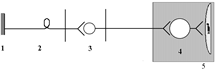 (ПСНС)
(ПСНС)
1 – рецептор; 2 – афферентный нейрон; 3 – центральный (преганглионарный) нейрон; 4 – ганглионарный нейрон (парасимпатический ганглий); 5 – эффектор (гладкая мышца).
12. Понятие о рефлексе. Строение рефлекторной дуги и функции его звеньев. Нарисовать схему рефлекторной дуги вегетативного рефлекса.(см. выше)
Рефлекс – ответная реакция организма на раздражение рецепторов, осуществляемая при обязательном участии нервной системы
Первое звено (рецептор) воспринимает раздражение, трансформируя энергию раздражения в нервный импульс. Второе звено (афферентный нейрон) проводит импульсы в ЦНС. Третье звено Вставочные нейроны – передают импульсы к эфферентному нейрону и обеспечивают связь данной рефлекторной дуги с другими отделами ЦНС. Четвертое звено (эфферентный нейрон) перерабатывает информацию, поступающую к нему от вставочных нейронов ЦНС и формирует ответ в виде нервных импульсов, посылаемых к 5-у звену – к рабочему органу.
13. Проприорецепторы, их виды, функции, механизмы возбуждения.
Проприорецепторы-рецепторы опорнодвигательного аппарата(мышц, сухожилий, фасций, суставов), что принимают участие во всех актах, что связаны с работой мышц, суставов и сухожилий.
 К проприорецепторам относят мешочные веретена и сухожильные рецепторы (Гольджи). Функции: принимают участие в осознание направления и скорости движения конечностей, определить положение отдельных частей тела в пространстве, построение трехмерного чувствительного окружающего мира(стереогноз).
К проприорецепторам относят мешочные веретена и сухожильные рецепторы (Гольджи). Функции: принимают участие в осознание направления и скорости движения конечностей, определить положение отдельных частей тела в пространстве, построение трехмерного чувствительного окружающего мира(стереогноз).
Мышечные веретена- расположены параллельно к мышечным волокнам, и при расслаблении мышцы, веретена растягиваются. Тем самым учащая ПД в афферентном нейторе возбуждают свой центр, тормозя нейроны центра антагониста
Рецепторы Гольджи- расположены между сухожильными волокнами мышц(последовательно), при напряжение мышцы, сухожилья растягиваются, деформируя рецептор, в следствии чего возрастает частота ПД в афферентном нейроне, что образует синапс с тормозными интернейронами, тем самым тормозит гомономные мотонейроты.
14.Возбуждение в ЦНС. Механизмы и закономерности передачи возбуждения в центральных синапсах.
Механізм передачі збудження через центральний аксосоматичний хімічний синапс полягає в наступному: ПД поширюється по мембрані аксона далі по мембрані пресинаптичній підвищення проникності пресинаптичної мембрани для іонів Сa2+ вхід їх в нервове закінчення за градієнтом концентрації вихід медіатора(АХ/НА) в синаптичну щілину дифузія медіатора до постсинаптичної мембрани взаємодія з мембранними циторецепторами(N-холино/адрено) збільшення проникності постсинаптичної мембрани для іонів Na+ вхід іонів Na+ в тіло клітини через постсинаптичну мембрану деполяризація мембрани (ЗПСП місцеве збудження) ЗПСП як місцеве збудження поширюється на сусудні ділянки постсинаптичної мембрани та мембрани аксонного горбика з допомогою місцевих струмів. Ці струми в незбуджених ділянках мембрани мають вихідний напрям, тому викликають деполяризацію мембрани. Цікавим є виникнення місцевих струмів між постсинаптичною мембраною (там ЗПСП) та мембраною аксонного горбика – початковий сегмент аксона, мембрана якого має найбільшу збудливість, поріг деполяризації (ΔЕ) там складає 10-15 мВ. Тому ПД виникає під впливом місцевих струмів саме там. Це відбувається, якщо під впливом місцевих струмів деполяризація мембрани аксонного горбика досягає критичного рівня виникнення серії ПД ритмічний розряд нейрона.
Особливості передачі збудження через центральні аксо-соматичні хімічні синапси.
1. Одностороннє проведення.
2. Сповільнення проведення – характеризується наявністю синаптичної затримки – час від виникнення ПД на пресинаптичній мембрані до виникнення ПД на мембрані аксонного горбика. Воно складає 2-3 мс.
3. Швидке порушення проведення через виснаження запасів медіатора.
Ці три особливості характерні для будь-яких хімічних синапсів, бо синаптична затримка та швидкість розвитку втоми в центральних синапсах більша, ніж в нервово-м’язових. Наступні дві особливості характерні лише для центральних синапсів.
4. Один ПД через центральний синапс не проходить тому, що зумовлює на постсинаптичній мембрані виникнення одного ЗПСП, який має амплітуду 1 мВ та тривалість 15 мс. Оскільки поріг деполяризації аксонного горбика складає 10-15 мВ, один ЗПСП, що поширився на мембрану аксонного горбика, не може викликати деполяризацію цієї мембрани до критичного рівня.
5. Умовою передачі збудження через центральний нервовий синапс є сумація ЗПСП на тілі нейрона.
До медіаторів, що викликають деполяризацію постсинаптичної мембрани та приймають участь в передачі збудження, відносять: ацетилхолін, норадреналін, серотонін та багато інших.
15.Торможение в ЦНС. Его виды и физиологическая роль. Механизмы и закономерности пре- и постсинаптического торможения. Нарисовать схемы пре- и постсинаптического торможения.
Торможение- нервный процесс, что обусловлен возбуждением и внешне проявляется угнетением другого возбуждения
| За локализацией | За характером поляризации | За строением тормозных нейронных цепей |
| -пресинаптическое -постсинаптическое | -гиперполяризационное -деполяризационное | -реципрокное -возвратное -латеральное |
Торможение является важным фактором координационной деятельности ЦНС, участвует в обработке информации, поступающей к нейрону, выполняет охранительную роль.
Пресинаптическое торможение-связано с угнетением проведения нервных импульсов аксональных(пресинаптических) окончаниях. К возбужденному аксону подходит вставочный тормозной аксон что выделяет тормозной медиатор ГАМК
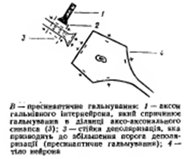 Постсинаптическое торможение- обусловлено выделением из пресинаптического окончания аксона тормозного медиатора, что понижает или тормозит возбудимостьсомы иди дендридов(аксо-сомальный, аско-дендритный синапс)
Постсинаптическое торможение- обусловлено выделением из пресинаптического окончания аксона тормозного медиатора, что понижает или тормозит возбудимостьсомы иди дендридов(аксо-сомальный, аско-дендритный синапс)
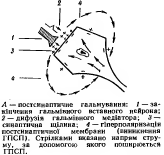
16. Суммация возбуждения и торможения нейронами ЦНС
Суммация возбуждающих постсинаптических потенциалов (ВПСП) или тормозных постсинаптических потенциалов (ТПСП) в нейронах нервного центра, в результате которых может возникнуть импульсное возбуждение.
Различают:
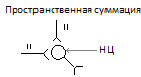
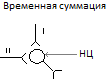
а)временную- при поступлении серии нервных импульсов по одному и тому же афферентному пути.
Б)пространственную-при которой импульсы подходят одновременно по многим афферентным волокнам.
17.Проводниковая функция СМ. Спинальный шок.

 Спинальный шок - резкое угнетение возбудимости и рефлекторной деятельности спинного мозга ниже места его травмы или перерезки. Возникает вследствие выключения активирующего влияния вышележащих отделов ЦНС на спинной мозг.
Спинальный шок - резкое угнетение возбудимости и рефлекторной деятельности спинного мозга ниже места его травмы или перерезки. Возникает вследствие выключения активирующего влияния вышележащих отделов ЦНС на спинной мозг.
18. Фазные и тонические рефлексы СМ, их рефлекторные дуги и физиологическое значение.
Фазные р-сы – однократное сгибание(разгибание) конечности при однократном раздражении кожи или мышечных рецепторов. (участвуют в акте ходьбы)
Тонические р-сы – р. что обеспечивают сохранение положения тела в пространстве, взаимное расположениеего частей, равновесие тела, конечностей и глаз с положением головы, включает статические и статокинетические рефлексы.
19.Двигательные рефлексы заднего и среднего мозга, децекебральная ригидность.
Сгибательные, разгибательные, ритмические, познотонические
Після операції децеребрації у тварини виникає децеребраційна ригідність, яка у кішки проявляється у слідуючому: передні і задні кінцівки розігнуті; голова закинута назад за рахунок розгинання шиї; хвіст піднятий; тулуб розігнутий. Ця поза кішки пов’язана з гіпертонусом розгиначів. Позу децеребраційної тварини (децеребраційна ригідність) можна охарактеризувати як посилену позу антигравітації. Згадаємо, що спинний мозок забезпечував формування м’язового тонусу рефлекторної природи, але не забезпечував формування пози антигравітації із-за малої сили тонусу
 20.РФ ствола мозга, механизм поддержания ее тонуса. Нисходящие и восходящие влияния РФ.
20.РФ ствола мозга, механизм поддержания ее тонуса. Нисходящие и восходящие влияния РФ.
РФ - скопление нейронов различных типов и размеров, связанных множеством волокон, идущих в различных направлениях и образующих сеть на всем протяжении ствола мозга, а также в шейных и верхнегрудных сегментах спинного мозга. Для поддержания тонуса РФ получает импульсы от всех рецепторов организма и от всех отделов ЦНС. Нейроны РФ являются полимодальными, посылают импульсы ко всем отделам ЦНС.
Аферентних шляхів РФ дуже багато, вона отримує інформацію від:
- усіх специфічних висхідних еферентних шляхів, які віддають колатералі до ядер РФ, заходячи в стовбур мозка;
- від іних утворень ЦНС, а саме:
-від кори головного мозку;
-від гіпоталамуса;
-від мозочка;
-від базальних ядер;
-від інших утворень.
Еферентні зв’язки РФ можна поділити на висхідні та низхідні.
Висхідні зв’язки РФ та висхідні впливи:
- через неспецифічні ядра таламуса передає інформацію всім відділам головного мозку та має неспецифічний активізуючий вплив на клітини. Цей вплив забезпечує бодьорий стан організму та визначає рівень активності клітин кори. РФ має окремі структури (гіпногенні зони), котрі зменшують ступінь (рівень) активності клітин кори та зумовлюють стан сну;
- по ретикулогіпоталамічним шляхам передає інформацію від різних рецепторів в гіпоталамус.
Характер низхідних ретикулоспінальних впливів:
- від медіального ядра РФ заднього мозку починається ретикулоспінальний (входить до складу МНС) – специфічно активує мотонейрони флексорів та гальмує мотонейрони екстензорів тулуба та проксимальних відділів кінцівок;
- від інших ядер РФ беруть початок шляхи, які неспецифічно активують чи гальмують спінальні мотонейрони і таким чином визначають ступінь їхньої активності;
- від вегетативних центрів РФ (дихальний, гемодинамічний) по ретикулоспінальним шляхам передається інформація до симпатичних рухових нейронів спинного мозку при регуляції відповідних функцій організму.
21. Роль среднего мозга в обеспечении положения тела. Двигательные рефлексы среднего мозга.
Середній мозок (СрМ) за участі сітчастої речовини опрацьовує аферентну інформацію, яка поступає в спинний та задній мозок. Нова інформація поступає в СрМ від зорових та слухових рецепторів. Її опрацювання відбувається в ядрах передніх (від зорових рецепторів) та нижніх (від слухових рецепторів) горбків. На основі опрацьовання інформації від усіх цих рецепторів СрМ здійснює контроль за станом зовнішнього та внутрішнього середовища організма.
Важливими надсегментарними руховими ядрами СрМ є:
1) червоні ядра – від них інформація від нейронів спинного мозку передається по шляхах що перехрещуються (руброспінальні шляхи – елемент ЛНС);
2) ретикулярна формація;
3) чорна субстанція – знаходиться в середньому мозку, але функціонує як єдине ціле з базальними ядрами.
Руброспінальні впливи характеризуються активацією - та -мотонейронів флексорів та гальмуванням - та -мотонейронів екстензорів. Ці впливи протилежні вестибулоспінальним. Завдяки руброспінальним впливам тонус м’язів мезенцефальної тварини більш рівномірніший, ніж у децереброваної (тонус флексорів та екстензорів більш збалансований). Тому, поза мезенцефальної тварини нормальна. Тобто, посилена антигравітаційна поза децереброваної тварини перетворюється в нормальну у мезенцефальної завдяки руброспінальним впливам.
За участю рухових ядер СрМ здійснюються позо-тонічні рефлекси двох видів:
1.Випрямні – забезпечують відновлення порушеної пози. У тварин, що мають шию, випрямні рефлекси мають 2 фази:
а) Рефлекс випрямлення голови – він починається з подразнення отолітових вестибулорецепторів у відповідь на порушення нормального положення голови в просторі (наприклад при падінні). Рівень замикання цього рефлексу – СрМ. Для його здійснення необхідна участь червоних ядер та РФ СрМ. По низхідних шляхах від цих структур інформація передається до мотонейронів спинного мозку перерозподіл тонусу м’язів шиї голова набуває нормальне положення в просторі (тім’ячком доверху).
б) Рефлекс випрямлення тулуба – починається з подразнення пропріорецепторів шиї, що спричиняється рефлексом випрямлення голови, так як при цьому відновлюється нормальне положення голови у просторі, але порушується її нормальне положення відносно тулуба. Вхід інформації від пропріорецепторів шиї здійснюється на рівні спинного мозку підйом до надсегментарних структур СрМ обробка цієї інформації передача до мотонейронів спинного мозку до м’язів, які забезпечують відновлення нормального положення тулуба відносно голови у просторі.
2. Стато-кінетичні – забезпечують збереження пози рівноваги при русі з прискоренням. При русі з лінійним прискоренням збуджуються отолітові рецептори, а при русі з кутовим прискоренням – рецептори півколових канальців. Вхід цієї інформації в ЦНС здійснюється на рівні заднього мозку підйом до структур СрМ обробка та передача низхідними шляхами до мотонейронів спинного мозку перерозподіл тонусу м’язів, який забезпечує збереження рівноваги під час руху з прискоренням. Важливим компонентом стато-кінетичних рефлексів є присінково-окорухові рефлекси ністагм очей більш продовжена фіксація предметів в полі зору під час руху краща орієнтація в просторі більш надійне збереження пози при русі.
Ще раз підкреслимо, що рівень замикання випрямних та стато-кінетичних рефлексів – СрМ. Тому, їх можливо отримати у мезенцефальної тварини і неможливо у децереброваної чи спінальної.
За рахунок обробки інформації від зорових та слухових рецепторів (за участю ядер чотирьох горбків) рухові ядра СрМ забезпечують здійснення орієнтаційних рефлексів – у відповідь на сильні звукові та світлові подразники мезенцефальна тварина повертає голову в бік подразника, насторожує вуха, в неї дещо посилюється тонус флексорів – тварина ніби готується до відповіді на цей подразник.
Роль СрМ в здійсненні фазичних реакцій залежить від рівня організації тварини:
- мезенцефальні земноводні (жаба) здатні до локомоцій – руху у просторі (стрибають, плавають);
- кішки та собаки не здатні до локомоцій, але здійснюють локомоторні синергії – злагоджені рухи кінцівок, що нагадують акт ходьби: їх інтегрують структури СрМ разом з структурами заднього та спинного мозку.
У людини СрМ не забезпечує формування нормальної антигравітаційної пози в спокої та підтримання її при русі. Для здійснення всіх цих функцій у людини необхідна участь кори великих півкуль (принцип кортикалізації функцій).
22.Мозжечок, его функции. Симптомы поражения.
Мозочок (Мз) відіграє дуже важливу роль в координації рухів і виконує цю функцію на основі своїх аферентних та еферентних зв’язків.
По аферентним волокнам сигнали поступають в кору мозочка, яка має складну будову та забезпечує тонку обробку поступившої інформації. Причому, кінцеву обробку інформації здійснюють грушоподібні клітини Пуркіньє – по відростках цих клітин (по їх аксонах) здійснюється еферентний вихід обробленої інформації з кори Мз. Далі, інформація здебільшого прямує до ядер Мз по аксонах їх нейронів здійснюється вихід інформації за межі Мз.
Аферентні зв’язки Мз:
1.Від рецепторів шкіри та пропріорецепторів Мз отримує інформацію по спіно-церебелярних шляхах; ця інформація забезпечує можливість контролю за рухом який здійснюється.
2.Від вестибулорецепторів (через вестибулярні ядра) – контроль за збереженням рівноваги при русі.
3.Від всіх рухових ядер стовбуру (ретикулярна формація, краєві ядра).
4.Від базальних ядер.
5.Від моторної і асоціативної кори (від моторної кори надходить копія команди до виконання руху, яка відправляється до мотонейронів; від асоціативної кори – інформація про ціль руху).
6.Від зорових та слухових рецепторів – контроль за рухами, які відбуваються.
Еферентні зв’язки Мз:
1.З руховими ядрами стовбуру (ретикулярна формація, вестибулярні ядра, червоні ядра), через які Мз здійснює вплив на мотонейрони і на м’язи.
2.З базальними ядрами.
3.З асоціативною та моторною корою
4.З гіпоталамусом.
Мз отримує інформацію про характер руху від рецепторів і при необхідності може швидко корегувати ці рухи та їх програму. При цьому різні відділи Мз здійснюють корекцію різних рухів.
Древній та старий Мз (флокуло-нодулярна система та хробак) – отримують інформацію від вестибулорецепторів, пропріорецепторів та шкірних рецепторів. Кора цих відділів передає інформацію до ядра шатра і далі до вестибулярного ядра до медіального ретикулярного ядра до мотонейронів, що іннервують м’язи тулуба та кінцівок.
Таким чином, кора древнього та старого Мз, отримуючи інформацію від пропріорецепторів, рецепторів шкіри про характер руху; від вестибулорецепторів – про збереження рівноваги, і при необхідності вносить корективи в грубі позо-тонічні рухи, чому передує передача інформації по шляхах МНС, які регулюють стан м’язів тулуба і проксимальних відділів кінцівок. Ця корекція дуже важлива для підтримання рівноваги (збереження пози) при русі.
Проміжна частина кори Мз, як і кора древнього та старого Мз, отримує інформацію про те, який рух здійснюється. Від рухової кори сюди поступає інформація про те, який рух повинен відбуватися (копія рухової програми). Кора проміжної частини Мз працює як блок порівняння – порівнює інформацію про те, який рух повинен відбуватися, і про те, який рух проходить. При неспівпаданні цієї інформації Мз може вносити такі корективи:
а)в рух, який здійснюється (виходячи на мотонейрони спинного мозку через стовбурові рухові ядра);
б)в рухову програму, передаючи інформацію по церебело-таламо-кортикальному шляху в рухову кору.
Вихід до рухової кори і до стовбурових рухових ядер із кори проміжної частини Мз здійснюється через вставне ядро.
За рахунок корекції рухів та програми Мз добивається їх повного співпадання (їх відповідності один одному).
Кора півкуль Мз пов’язана з базальними гангліями і з асоціативною корою. Завдяки цим зв’язкам вони формують програми рухів і Мз через зубчасте ядро передає їх в рухову кору для виконання.
Основні симптоми ураження Мз у людини. Мехпнізми їх виникнення.
1.Дистонія – порушення тонусу м’язів, що спричиняється порушенням впливу Мз на стовбурові рухові ядра (ретикулярна формація, вестибулярні ядра) на мотонейрони спинного мозку на м’язи тулуба та проксимальних відділів кінцівок.
2.Дизеквілібрія – перушення рівноваги. Пов’язана з випаданням коригуючих впливів Мз на вестибулярні ядра Дейтерса.
3.Атаксія – невпорядкованість рухів. Проявляється у вигляді:
а) дисметрія – порушення амплітуди рухів;
б) порушення направленості рухів.
В основі атаксії лежать порушення координації м’язів агоністів та антагоністів.
4.Асинергія – порушення злагодженості рухів. В основі її лежить порушення координації скорочення м’язів-агоністів.
5.Астенія – швидка втомлюваність. Вона пов’язана з виконанням великої кількості “зайвих” рухів для досягнення мети.
6.Адіадохокінез – порушення синхронного виконання рухів, іх послідовності та швидкості.
7.Дисметрія – порушення рівномірності і амплітуди рухів.
8.Дисартрія – порушення артикуляції мови.
23.Таламус, его функции
Таламус є структурою проміжного мозку і виконує, головним чином, сенсорні функції – за виключенням нюхового, в тамусі переключаються всі сенсорні шляхи, які йдуть в КГМ.
В таламусі є біля 40 ядер, які розрізняються по своїм функціям. Серед них виділяють:
1. Сенсорні перемикаючі (специфічні) ядра – вони отримують інформацію від специфічних сенсорних шляхів, переробляють її і передають в сенсорні зони КГМ. Разом з цими зонами специфічні сенсорні перемикаючі ядра таламуса беруть участь в формуванні відчуттів.
2.Неспецифічні – вони отримують інформацію від ретикулярної формації стовбура мозку по шляхах больової чутливості. Вони передають інформацію до всіх зон КГМ, здійснюючи на неї неспецифічний активуючий вплив.
3.Асоціативні – отримують інформацію від специфічних сенсорних перемикаючих ядер і від неспецифічних ядер таламуса. Вони передають інформацію в асоціативні зони КГМ (фронтальна, тім’яна, вискова). Разом з даними ділянками кори, асоціативні ядра таламуса приймають участь в розпізнаванні образів.
4.Неспецифічні перемикаючі:
а)лімбічні – отримують інформацію від гіпоталамуса та передають її в лімбічну кору. Вони беруть участь у формуванні емоцій та мотивацій.
б)рухові – отримують інформацію від базальних гангліїв, мозочка, стовбурових рухових ядер і передають її в рухову кору. Беруть участь в забезпеченні рухових реакцій організму.
24. Базальные ядра,их функции, симптомы поражения.
Базальні ядра знаходяться в глибині кінцевого мозку. До них відносяться:
- хвостате ядро;
- шкаралупа (разом з попереднім утворює смугасте тіло);
- бліда куля.
Як єдине ціле з базальними ядрами функціонують чорна субстанція та субталамічне ядро.
Ці ядра об’єднані між собою двосторонніми зв’язками, отримують інформацію від кори (асоціативних та рухових зон) та мозочка. Після відповідної обробки інформація від базальних ядер передається:
- через моторні (передні) ядра таламуса до рухової кори;
- через стовбурові рухові ядра (червоне ядро, вестибулярне ядро, ретикулярна формація) до мотонейронів спинного мозку до м’язів.
 В діяльності базальних ядер важливу роль відіграє рух збудження, який має назву циклу шкаралупи та циклу хвостатого ядра.
В діяльності базальних ядер важливу роль відіграє рух збудження, який має назву циклу шкаралупи та циклу хвостатого ядра.
1. Цикл шкаралупи – забезпечує участь базальних ядер в реалізації програм складних набутих рухових реакцій. Цикл починається з передачі інформації від премоторної зони кори (ПМК) до шкаралупи далі до блідої кулі (БК) через чорну субстанцію (ЧС) та субталамічне ядро (СТЯ) інформація, перероблена в базальних ядрах передається в першу моторну зону кори (МК) через моторні ядра таламуса.
Цикл хвостатого ядра – забезпечує участь базальних ядер в формуванні програм складних рухових реакцій. При здійсненні цього циклу інформація від асоціативних зон кори (АЗК) про те, яким повинен бути результат рухової реакції, направляється в хвостате ядро (ХЯ) далі в шкаралупу (Ш) і бліду кулю (БК) і далі через моторні ядра таламуса в рухову кору. Завдяки цьому рухова кора отримує готову рухову програму, яка підлягає наступній реалізації.
Необхідно пам’ятати, що у створенні програм рухових актів окрім базальних ядер важливу роль відіграє кора мозочка та його зубчасті ядра.
Симптоми ураження базальних ядер:
1.При ураженні смугастого тіла:
-гіпотонія м’язів;
-гіперкінези, в тому числі у спокої;
2.При ураженні блідої кулі та чорної субстанції:
-гіпокінезія, рухи втрачають індивідуальне забарвлення за рахунок зникнення міміки та пантоміміки;
-воскова ригідність – людина може довго утримувати ненормальну (незручну) позу, положення кінцівок;
-тремор спокою.
25Понятие о лимбической системе. Гипоталямус его связи и функции.
До складу лімбічної системи входять:
1.Лімбічна (стародавня і стара) кора:
-гіпокамп;
-гіпокампова закрутка (звивина);
-поясна закрутка;
-нюховий мозок (нюхові цибулини, горбки, ділянки кори над мигдалинами).
2.Підкіркові утворення:
-мигдалеподібний комплекс;
-лімбічні ядра таламуса;
-ядра перегородки;
-гіпоталамус;
3.Лімбічна частина ретикулярної формації середнього мозку.
Структури ЛС пов’язані між собою чисельними кільцевими зв’язками, що забезпечує можливість тривалої рециркуляції збудження в системі.
ЛС отримує інформацію практично від усіх рецепторів організму за рахунок аферентних зв’язків з:
-лімбічних ядер таламуса;
-ретикулярної формації середнього мозку;
-чисельних рецепторів гіпоталамуса (осмо-, термо-, глюко-, тощо).
Всі ці структури отримують інформацію від усіх рецепторів, окрім нюхових.
Такий обширний аферентний вхід дозволяє ЛС контролювати стан організму і ситуацію, в якій він знаходиться.
Лімбічна система забезпечує:
1.Формування мотивацій – стан організму, що виникає на базі потреби і змушує здійснювати реакцію, спрямовану на задоволення даної потреби.
2.Формування емоцій – стан організму, що виникає на базі потреби (з урахуванням можливості її задоволення) і мобілізують можливості організму для задоволення даної потреби, а також дозволяють швидко оцінювати інформацію, що надходить з точки зору її значення для задоволення даної потреби.
26.Сенсорные, ассоциативные и моторные зоны коры ГМ и их функции.
Сенсорні, асоціативні, моторні зони кори формують нову кору – неокортекс.
Сенсорні зони кори відповідають представництву окремих сенсорних систем (аналізаторів) у різних ділянках кори.
Так, кіркове представництво зорового аналізатора локалізується у потиличній зоні кори (шпорна закрутка), слухового – у висковій зоні, соматосенсорного – у постцентральній закрутці.
Сенсорні зони кори, діючи разом з іншими елементами специфічних каналів зв’язку в сенсорних системах, забезпечують формування відчуттів.
Асоціативні зони кори – лобна, тім’яна, вискова забезпечують виконання вищих кіркових функцій, а саме:
1.Розпізнавання образів (разом з асоціативними ядрами таламуса);
2.Здійснення абстрактного мислення, мови;
3.Беруть участь у формуванні пам’яті
4.Забезпечують вибір цілей рухових реакцій та на основі цього приймають участь у формуванні та виборі конкретних програм цих рухів.
5.Приймають участь у формуванні, зберіганні та реализації вроджених та набутих програм рухових реакцій організму.
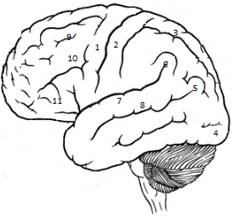
Моторні зони – їх декілька. Головна моторна зона знаходиться у прецентральній звивині, премоторна зона – попереду головної (первинної) моторної зони і в глибині сільвієвої борозни. Моторні нейрони ї також у зоні представництва соматосенсорного аналізатора – постцентральна закрутка.
1-предцентральная/движение
2-постцентральная/ чувствительность
3-верхняя теменная/стереогноз
4-затылочная/зрение
5-угловая/чтение
6-краевая/праксия
7-верхняя височная/слуховой анализатор
8-центр Вернике(пониманае речи)
9-письмо
10-поворот глаз противоположно повороту головы
11-центр Брока(артикуляция речи)
27.Электрическая активность ГМ, частотно- амплитудная характеристика основных ритмов ЭЭГ. Клиническое значение ЭЭГ.
Электроэнцефалография - регистрация суммарной электрической активности головного мозга с поверхности кожи головы. ЭЭГ – кривая записи суммарной электрической активности головного мозга: альфа-, бета-, тэта- и дельта-волны.
Альфа-волны. Частота 8 – 13 Гц, амплитуда 25 – 50 мкВ. Закрыты глаза
Бета-волны. Частота 14 – 30 Гц, амплитуда 20 – 25 мкВ. Открыты глаза
Тэта-волны Частота 4 –7 Гц, амплитуда 100 –300 мкВ. Переход к сну
Дельта-волны Частота 0.5 – 3.5 Гц, амплитуда 100 –300 мкВ глубокий сон
28.Структурно-функциональная организация СНС: НЦ, медиаторы, специальные рецепторы, вторичные мессенджеры.
НЦ- nuc. Intermedius lateralis C8 – L2-3
Преганглионарные: медиаторы – АХ (аценилхолинэстераза)
Рецепторы – Н-холинорецепторы
Постганглионарные: медиаторы – А, НА, ГАМК, серотонин(моноаминооксидаза)
Рецепторы:
- α1-гладкие мышцы(сужение сосудов, сокращение миометрия)
- α2- (снижает выделение медиаторов)
- β1-сердце (^ ЧCC)
- β2-гладкая мускулатура(расслабляет коронарные артерии, а. бронхов, ГМ, мочевого) Курареподобные в-ва/ бензогексоний
29.Влияние СНС на функции организма
Активація симпатичної нервової системи в організмі відбувається у стані напруження, коли необхідно забезпечити мобілізацію функцій для вирішення важливої задачі (фізична робота, емоційний стрес). Симпатична нервова система забезпечує ерготропну перебудову функцій організму, а саме:
- збільшує частоту та силу серцевих скорочень;
- звужує більшість судин (такі впливи на серце і судини призводить до збільшення системного артеріального тиску – САД);
- розширює бронхи створюються умови для оптимальної вентиляції легень;
- пригнічує секреторну та рухову активність органів травної системи (функції цієї системи в стані напруження є вторинними);
- збільшує дієздатність скелетних м’язів (через зміну обміну речовин та кровотоку в м’язах);
- стимулює виділення гормонів мозкової речовини наднирників;
- збільшує збудливість рецепторів і центрів, дієздатність клітин кори головного мозку;
- змінює обмін речовин в організмі (стимулює глікогеноліз і ліполіз розпад глікогену та жирів збільшення концентрації в крові глюкози та жирних кислот субстратне забезпечення посиленого функціонування скелетних м’язів і клітин ЦНС).
-расширяет зрачок.
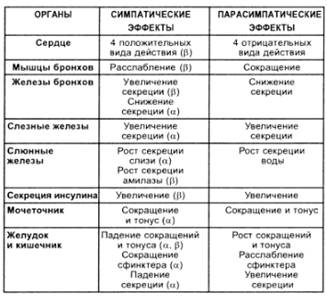
30. Строение ПСНС: синапсы, медиаторы, циторецепторы, блокаторы.
НЦ- nuc. Intermedius medialis S2-3(на границе передних и задних рогов)
Кранио-бульбарный(3,7,9,10)
Преганглионарные: медиаторы – АХ (аценилхолинэстераза)
Рецепторы – Н-холинорецепторы
Постганглионарные: медиаторы – АХ (аценилхолинэстераза)
Рецепторы: -М- холинорецепторы
Курареподобные в-ва / бензогексоний/ атропин
31. Влияние ПСНС на функции организма.
Парасимпатична нервова система активується після стану напруження і забезпечує трофотропну перебудову функцій організму, завдяки якій проходить відновлення пластичних та енергетичних ресурсів організму, що витратились під час стану напруження.
Вплив парасимпатичної нервової системи:
- зменшення ЧСС перехід серця на режим економної діяльності активний перебіг відновлювальних процесів в міокарді;
- звуження бронхів;
- активація секреторної і рухової активності органів травної системи сприятливі умови для перетравлювання та всмоктування поживних речовин посилення відновлювальних процесів;
- посилення секреції інсуліна посилення утилізації глюкози клітинами посилення відновлювальних процесів.
32.Роль центров СМ, продолговатого, среднего мозга в регуляции вегететивной функции.
Спинний мозок. Первинні симпатичні та парасимпатичні центри бокових рогів спинного мозку забезпечують реалізацію елементарних рефлексів. Парасимпатичні рефлекси (дефекація, сечовипускання, статеві) у людини в значному ступені контролюються корою головного мозку. Реалізація симпатичних спінальних рефлексів (судинно-рухових, потовидільних) перебуває під контролем сітчастої речовини заднього мозку.
Задній мозок. Вміщує в собі первинні парасимпатичні центри у вигляді відповідних ядер VII, ІX (регуляція слиновиділення) та X пар ЧМН (регуляція рухової та секреторної функції органів травної системи, роботу серця, тонус бронхіальної мускулатури). Окрім того, структури сітчастої речовини заднього мозку утворюють суднно-руховий, кардіостимулюючий та дихальний центри. Саме тому, роль заднього мозку в регуляції вегетативних функцій дуже велика. Цей рівень є першим, що може забезпечити зміну вегетативних функцій в інтересах цілісного організму (наприклад, при фізичній роботі) тут вже розміщені інтегративні центри, як приклад – сітчаста речовина.
Середній мозок. Вміщує в собі вегетативне ядро ІІІ пари чмн (ядро Якубовича), за участю якого здійснюються зіничний рефлекс (звуження зіниці) та рефлекс акомодації (зміна кривизни кришталика). Тобто, цей центр приймає участь в забезпеченні умов для ясного (чіткого) бачення.
33. Роль гипоталамуса, лимбической системы, неокортекса в регуляции вегететивных функций.
Гіпоталамус – структура проміжного мозку. Він виконує важливі функції завдяки наявності в ньому таких особливих структур:
1. В його склад входять інтегративні центри, що контролюють стан (та інтегрують):
- симпатичних центрів, а саме, сітчаста речовина (це здійснюють переважно ядра заднього гіпоталамуса);
- первинних парасимпатичних центрів стовбура мозку та спинного мозку. Саме тому гіпоталамус інтегрує нервові механізми регуляції вегетативних функцій (симпатичні та парасимпатичні) між собою.
2. До складу медіального гіпоталамуса входять нейросекреторні клітини, що обробляють інформацію як будь-які інші нервові клітини (шляхом сумації збуджень та гальмувань), та виділяють із нервових закінчень не медіатори (час життя, яких короткий, бо вони швидко руйнуються відповідними ферментами), а більш стійкі гормони:
а) Ліберини та статини, які по системі ворітних судин поступають в аденогіпофіз (передня та середня долі) і стимулюють (ліберини) чи гальмують (статини) виділення відповідних гормонів:
| Гормони гіпофіза. | Гормони гіпоталамуса. | |
| Ліберини. | Статини. | |
| Соматотропний | Соматоліберин. | Соматостатин. |
| Тиреотропний. | Тиреоліберин. | --- |
| Адренокорти. | Кортиколіберин. | --- |
| Гонадотропний. | Гонадоліберин. | --- |
| Пролактин. | Пролактоліберин. | Пролактостатин. |
| Меланоцито-ст. | Меланоліберин. | Меланостатин. |
б) Нейросекреторні клітини супраоптичного та паравентрикулярного ядер виділяють гормони вазопресин та окситоцин, які по аксонах цих нейронів надходять в нейрогіпофіз (задня доля), а звідти вже виділяються в кров.
Тропні гормони аденогіпофіза (тиреотропний, адренокортикотропний, гонадотропний) стимулюють виділення відповідних гормонів ендокринними залозами організму (щитовидною, кірковим шаром наднирників, статевими).
Тому, гіпоталамус виступає структурою, що інтегрують гуморальні (ендокринні) механізми регуляції вегетативних функцій.
Наявність в гіпоталамусі центрів, що інтегрують нервові мехінізми регуляції вегетативних функцій, а також нейросекреторних клітин, дають йому змогу інтегрувати нервові та гуморальні механізми регуляції вегетативних функцій.
Лімбічна кора (стара та древня) приймає участь в регуляції вегетативних функцій при формуванні мотивацій та емоцій.
Нова кора за участю лімбічної системи та гіпоталамуса бере участь в регуляції вегетативних функцій організму при здійсненні різноманітних реакцій поведінки.
Сенсорные с-мы и ВНД
34. Общие пинципы структурной и функциональной организации анализаторов.
Анализатор- совокупность нервных образований, воспринимающих и анализирующих изменения внешней и внутренней среды организма.Выделяют: Внешние и внутренние анализаторы, положения тела и болевой анализатор.
Его структурные элементы.
1) Периферический отдел – рецепторы;
2) проводниковый – афферентные нейроны и проводящие пути;
3) корковый отдел – проекционная и ассоциативная зоны коры больших полушарий.
35. Рецепторы, их классификация, механизмы возбуждения.
Рецептор служит для восприятия раздражений и их первичного анализа. Восприятие раздражения осуществляется путем трансформации энергии раздражения в нервный импульс.Различают первичные и вторичные рецепторы. В первичном рецепторе воспринимающая зона представляет собой окончание дендрита чувствительного нейрона; во вторичном рецепторе воспринимающей зоной является специальная рецепторная клетка, синаптически связанная с окончанием дендрита чувствительного нейрона. Первичные: все кожные рецепторы и рецепторы внутренних органов, проприорецепторы, рецепторы обоняния, термо- и хеморецепторы ЦНС. Вторичные: вкусовые, фоно-, фото-, вестибулорецепторы. В первичных рецепторах – рецепторный потенциал – потенциал действия. Во вторичных – рецепторный потенциал – выделение медиатора – генераторный потенциал – потенциал действия.
36.Структурно-функциональная организация соматосенсорного анализатора.
Осязание – это совокупность ощущений, возникающих при раздражении рецепторов кожи, слизистых (прикосновение, давление, вибрация, щекотка, температура, боль) и рецепторов опорно-двигательного аппарата. Перерабатывает сенсорную информацию нескольких модальностей, получая ее от кожи, мышц, суставов, внутренних органов и кровеносных сосудов
-тактильная чувствительность(чувство осязания)
А.свободные нервные окончания(волосяных фоликулов)- механорецепторы, (кожи)-терморецепторы,
Б.инкапсулированные нервные окончания(тельца Пачини, мейснера)- давление, (Руффина, Пинкуса-Игго,диски меркеле)- тактильная, (колбы Краузе)- вибрация
По скорости адаптации к раздражению: -быстроадаптирующиеся(Мейснера, Пачини) -медленно(Руффи, Меркеля)
-проприоцептивная(сухожильные рецепторы Гольджи, мышечные волокна, механорецепторы суставной капсулы)
-терморецепторы.
Центральные отростки первичных сенсорных нейронов - >в составе задних корешков входят в СМ и в задних рогах разделяются на контрлатеральные, до переключательных ядер заднего столба продолговатого мозга(информ. Попадает на противоположную сторону по Лемнистскому пути) - > к проэкционным ядрам таламуса и затем в постцентральную извилину(соматосенсорная кора)
37.Современные представления о механизме болевой чувствительности. Физиологические основы наркоза и обезболивания.
Болевые ощущения возникают после раздражения болевых (специализированных свободных нервных окончаний) и любых других рецепторов при действии любого сильного раздражителя. При ощущении чувства боли повышается мышечный тонус, усиливается сердечная деятельность и дыхание, угнетается функция пищеварительного тракта и почек.
Болевые рецепторы полимодальные (механо, термо,хемо) Центральный орган ноцицепции гипоталямус (латеральные ядра)
Центральные отростки первичных сенсорных нейронов - >в составе заднего корешка заходит в СМ, где располагается синапс с нейронами второго порядка, что переходят на противоположную сторону, где образуют спиноталямический(к нижнему заднему ядру таламуса) и спиноретекулярный тракты -> таламус затем далее на постцентральную извилину(соматосенсорную кору).
Анальгизирующим действием обладают опиатные пептиды (дайнорфины, эндорфины) синтезируемые гипоталамусом.
Местная анестезия – блокада болевой импульсации переферических нервных волокон(блокируют Na каналы)Новокаин, лидокаин.
Общая анестезия – ингаляционная, в/в, сегментарная, основана на введение наркотических вещ-в что блокируют проведение болевых импульсов в кору ГМ, и гипоталамус.
38. Структурно-функциональная организация зительного анализатора. Механизм аккомодации.
1Фоторецепторы (палочки – родопсин/ светоощущение, колбочки – йодопсин/цветоощущение)
Родопсин - > метародопсин= ретиналь(витамин А) + опсин (848 АК)
Под действием света родопсин распадаясь изменяет МП фоторецепторов. Скототопическая с-ма(палочки)
2Биполярные клетки 3Ганглиозные клетки чьи аксоны образуют зрительный нерв, затем зрительный перекрест и далее зрительный тракт средний мозг(зрачковый рефлекс), верхние бугры четверохолмия (ориентировочный рефлекс), латеральные коленчатые тела. Затылочные доли.
Аккомодация – механизм, с помощью которого глаз настраивается на рассмотрение удаленных или близких предметов(за счет изменения кривизны хрусталика), зависит от степени натяжения капсулы хрусталика, что циновой связкой прикреплена к ресничному телу.
Сокращение мышцы - >раслабление циновой связки - >выпуклый хрусталик - >вблизи(70,5 диоптрий) конвекция
Расслабление мышцы - >напряжение связки - >хрусталик уплощается - >вдаль
(58,6 диоптрий) дивергенция
39.Слуховая сенсорная система, ее строение и функции.
40. Структурно-функциональная организация вкусового анализатора
Анализ вкусовых свойств пищи осуществляется вкусовой сенсорной системой. На поверхности языка можно выделить зоны специфической чувствительности: кончик – сладкого, корень – горького, края – кислое, тело – соленое.
Рецепторы- вкусовые сосочки(с вкусовыми почками)
Проводники: ganglion geniculi(7 передние 2/3), ganglion inferius (9 задняя 1/3, 10) затем в составе nervus intermedius идут в мост - > продолговатый мозг - >
Центральный отдел: к коре к крючку и парагипокампальной извилине.
41. Структурно-функциональная организация обонятельного анализатора
Рецепторы: в обонятельной зоне(верхний носовой ход) находятся рецепторые
обонятельные клетки, на поверхности каждой обонятельной клетки имеется сферическое утолщение – обонятельная булава(с 6 -12 микроворсинками)
Проводник: располагается в bulbus olfactorius, дендриты которого в составе filae olfactorii взаимодействуют с обонятельными рецепторами, аксоны в составе tractus olfactorii следуют в центральный отдел.
Центральный отдел: распологается в коре в парагипокампальной извилине.
42.Функциональная асимметрия полушарий головного мозга. Роль коры в обеспечении высших психических функций человека.
Межполушарная асиметрия – неравнозначность функций правого и левого полушарий головного мозга.
Левое полушарие – логическое мышление, правое – интуитивное.
Латеризация мозга – обнаружена при обследовании больных эпилепсией, которым с целью лечения перерезали мозолистое тело. При письме задействовано преймущественно левое полушарие(правое при написание иероглифов) в формирование речи участвует только левое полушарие.
Принадлежность к левшам или правшам запраграмированна наследственно.
Увеличение роли коры большого мозга в регуляции функций и поведения организма в процессе филогенеза. У лягушки значительных изменений не наблюдается после полного удаления полушарий большого мозга. У голубя сохраняются только врожденные рефлексы на сильные раздражители; в покое сидит неподвижно, подброшенный вверх летит, но препятствий не облетает, не может самостоятельно питаться.
43. Значение работ И.М. Сеченова и И.П. Павлова в изучении ВНД. Отличие условных рефлексов от безусловных, их биологическая роль.
М. Сеченов распространил представления о рефлексе на психическую деятельность. И. П. Павлов открыл явление условного рефлекса и использовал его как метод исследования деятельности коры большого мозга, с помощью которого разработал основы учения о ВНД. открыл и изучил условные рефлексы, изучил типы ВНД, сформулировал понятия первая и вторая сигнальные системы, открыл закономерности иррадиации и концентрации процессов возбуждения в коре большого мозга и торможения условных рефлексов. И не требуют для своего возникновения специальных условий.
Условный рефлекс - приобретенная в онтогенезе реакция организма на раздражитель, ранее индифферентный для этой реакции (ранее ее не вызывавший) формируется у человека и высших животных при обезательном участие коры ГМ на базе безусловных рефлексов.
| Безусловные р-сы | Условные р-сы |
| 1. Врожденные наследственно передающиеся реакции,начинают функционировать после рождения. | 1.Реакции преобретенные в процесе индивидуальной жизни. |
| 2.являются видовыми (свойственны всему виду) | 2.индивидуальны |
| 3.постоянны, сохраняются в течение всей жизни | 3.непостоянны могут возникать, изменятся, исчезать. |
| 4.осуществляются за счет низших отделов ЦНС | 4. Являются преимущественно функцией коры ГМ |
| 5.Возникают в ответ на адекватные раздражители, действующие на определенное рецептивное поле. | 5.возникают на любые раздражители, на различные рецептивные поля. |
Условный рефлекс высшего порядка это условный рефлекс выработанный на базе другого условного рефлекса. Например, после выработки прочного оборонительного условного рефлекса на звонок включают другой сигнальный раздражитель – свет, а в качестве подкрепления дают звонок. В результате врабатывается оборонительный условный рефлекс второго порядка на свет.
Разновидности: 1) Простые – на одиночный раздражитель; 2) комплексные – один условный рефлекс на комплекс одновременно действующих раздражителей или несколько последовательно включаемых раздражителей; 3) цепные – несколько условных рефлексов на цепь раздражителей, каждый из которых вызывает другой условный рефлекс.
основные правила выработки условных рефлексов.
1) Неоднократное сочетанное действие двух раздражителей (индифферентного – сигнального, т.е. условного и подкрепления); 2) сигнальный раздражитель должен быть слабее и предшествовать подкреплению; 3) отсутствие посторонних раздражителей, вызывающих болевую реакцию или ориентировочный рефлекс; 4) здоровое бодрствующее животное.
44.Торможение условных рефлексов, виды коркового торможения. Физиологическое значение.
Различают:
Внешнее(при действии черезмерного БР, или в силу доминирования другого НЦ)
Внутреннее: - угасательное(предявление условного раздражителя без сочетания с БР, подкрепления)
- запаздывающее(задержка УР в следствии удленения интервала между условным раздражителем и БР, подкреплением)
-дифференцировочное(исчезновение УР на раздражители близкие к условно рефлекторным, в следствии не подкрепления БР, для уточнения условий р-са(Гц звуки))
- условное торможение (исчезновение УР на положительный условный раздражитель, если он предявляется в комплексе с новым У раздражителем, без БР))
Благодаря торможению уточняются условные рефлексы в соответствии к изменившимся условиям или происходит их временная отмена, если условный раздражитель утратит свое сигнальное значение.
45.Структура поведения с точки зрения теории функциональных систем П.К.Анохина. Потребности и мотивация, их роль в формировании поведения.
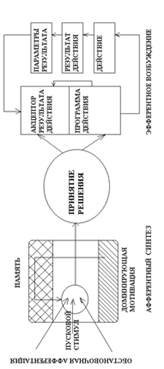
Функциональная система по П. К. Анохину – динамическая совокупность различных органов и систем организма, формирующаяся для достижения полезного (приспособительного) результата.
Потребность – источник активности организма, побуждение и цель его поведения в окружающем мире.
Формы потребностей: Биологические (потребность в пище, одежде и др.), социальные (потребность в социальном признании), духовные
 Мотивация - побуждение организма к действию. Причина возникновения мотиваций – наличие потребности или действие неблагоприятных факторов. Значение мотиваций – удовлетворение потребности и избегание или избавление от действия неблагоприятных факторов.
Мотивация - побуждение организма к действию. Причина возникновения мотиваций – наличие потребности или действие неблагоприятных факторов. Значение мотиваций – удовлетворение потребности и избегание или избавление от действия неблагоприятных факторов.
Основные компоненты функциональной системы 1) афферентный синтез; 2) принятие решения с одновременным формированием программы действия (3) и акцептора результата действия АРД (4); 5) действие; 6) параметры результата действия; 7) обратная афферентация от рецепторов результата действия к АРД.
46.Эмоции, механизм их возникновения и биологическая роль.Медицинские аспекты отрицательных эмоций.
Эмоция – реакция организма на внешние или внутренние раздражители, сопровождаемая ярко выраженными переживаниями. Лимбическая система (Гиппокамп, поясная извилина, гипоталамус, свод, миндалина, таламус) является морфологическим субстратом для эмоциональных реакций.
Основные факторы (причины), вызывающие отрицательные и положительные эмоции.
Положительные – удовлетворение потребностей, известие о приятном событии. Отрицательные – невозможность удовлетворения потребностей, известие о неприятном событии.
Основные компоненты эмоциональной реакции: 1) Двигательные и голосовые реакции, 2) вегетативные (изменение интенсивности функционирования внутренних органов и желез внутренней секреции), 3) интроспективные (изменение состояния внутреннего мира человека (его "я") и восприятия внешнего реального мира).
Изменение ритма сердцебиений, артериального давления, дыхания, сужение или расширение зрачка, пилоэрекция, изменение уровня саливации, гормональные сдвиги основные вегетативные компоненты, сопровождающие выраженную эмоциональную реакцию
стадии развития эмоционального состояния (по Г. И. Косицкому).
Стадии развития эмоционального состояния: Стадия напряжения(Состояние внимания, мобилизации, активности (ВМА). Повышается работоспособность, усиливается функция органов и систем, обеспечивающих решение данной задачи), стадия стенической отрицательной эмоции(Повышение эмоционального напряжения, что проявляется состоянием ярости (гнев, негодование) и сопровождается предельным усилением активности органов и систем. Достигается максимальная мобилизация ресурсов организма для выполнения стоящей перед ним задачи), стадия астенической отрицательной эмоции(Возникает при длительном эмоциональном напряжении, когда истощаются интеллектуальные и энергетические резервы организма. Подавляется умственная и физическая активность, развивается состояние "ужаса, страха, тоски".), стадия невроза(Развитие невротического состояния, "переход" в болезнь. Применение сверхсильных или необычных чрезвычайных раздражителей, выработка тонких дифференцировок, частые нарушения динамического стереотипа, быстрая смена тормозного и возбуждающего раздражителей ("ошибка"). Положительные эмоции оказывают благотворное влияние на состояние здоровья человека и стимулируют физическую и умственную деятельность; отрицательные стенические эмоции мобилизуют физические ресурсы и умственную деятельность для достижения цели, но чрезмерно выраженные или часто повторяющиеся оказывают негативное влияние на здоровье организма.
47.Типы и особенности ВНД человека.Основные характеристики нервных процессов, определяющ тип ВНД.
Сильный, уравновешенный, подвижный (сангвиник) «живой»;
сильный, уравновешенный, инертный (флегматик) «спокойный»;
сильный, неуравновешенный, подвижный (холерик) «взрывной» «безудержный»;
слабый, неуравновешенный, инертный (меланхолик) «аранжирейный».
Сила нервных процессов – при измерение силы процессов возбуждения используют кривую зависимотси величины условной реакции от силы раздражителя. Сила условной реакции перестает увеличиватся при определенной интенсивности условного сигнала.(эта граница и характеризует силу нервных процессов)
Уравновешенность нервных процессов – сравниваются процессы возбуждения и торможения у данного животного.Взаимокомпенсированны – уравновешенный, при преобладании процессов возбуждения – неуравновешенный «взрывной», при преобладании процессов торможения – неуравновешенный «аранжерейный»
Подвижность нервных процессов – скорость перехода положительных условных рефлексов в тормозные и обратно (степень лабильности).
48.Первая и вторая сигнальная система. Симптомы нарушения речи.
Первая сигнальная система – это система условнорефлекторних связей, что формируется в коре полушарий ГМ животных и человека под влиянием безусловных раздражителей.
Вторая сигнальная система – свойственна человеку, система условнорефлекторных связей, что формирует в высших отделах ЦНС под влиянием речевых сигналов.
Симптомы нарушения речи: - моторная афазия(х_х центра Брока)не может говорить,
- сенсорная афазия (х_х центра Вернике) не понимает слов,
- затылочная афазия((х_х угловой извилины) забывание слов, невозможность их чтение
49.Память, классификация, механизмы. Физиологическая роль пептидов в регуляции памяти, обучения.
Память – способность мозга человека преобретать и сохранять полученную в процессе индивидуальной жизни информацию, а также неоднократно использовать эту информацию в дальнейшем(воспроизводить). Энграмма – структурно-химический или электрофизиологический след памяти.
Виды памяти: сенсорная(0.1 – 0.5 сек), коротковременная (5 -60 сек до 10 мин), промежуточная (от мин до 3 недель), долговременная (возможна на протяжение всей жизни)
Так же различают: логическую, образную, зрительную, тактильную, моторную,обонятельную виды памяти.
Мгновенная (сенсорная) память соответствует ощущениям сразу после выключения сигнала; кратковременная – удержание информации в сознании и после исчезновения ощущений от действующего сигнала. В обоих случаях – это последействие в рецепторах и нейронных сетях (электрохимические процессы).
Основные изменения в нейронах при переходе кратковременной памяти в долговременную (консолидация памяти).
Химические и структурные изменения в нейронах обеспечиваются электрическими процессами. Сначала в синаптических структурах возрастает концентрация кальция, затем – число рецепторов (например, глютаматных, холинорецепторов), увеличивается синтез белков в нейронах.
В мозгу активно обучающихся животных содержится больше РНК; инъекция РНК ускоряет запоминание; стимуляция выработки РНК в организме (например, монофосфатом нуклеотидов, оротовой кислотой) также улучшает запоминание
Гиппокамп играет особо важную роль в процессах консолидации памяти, височная доля, миндалевидный комплекс. Например, одновременное удаление гиппокампа и миндаливидных тела исключает возможность дальнейшего обучения. Ранее закодированная в мозге информация сохраняется
емкость мгновенной – безгранична; емкость кратковременной – ограниченное число цифр, букв, предметов (обычно несколько); емкость долговременной – практически безгранична, зависит от значимости запоминаемой информации и ее систематизации.
50.Сон, его виды, фазы, механизмы.
Сон – периодически появляющаяся особая активность головного мозга, при которой выключены сознание и механизмы поддержания естественной позы, снижена чувствительность анализаторов.
Основные фазы сна по характеру ЭЭГ "Медленноволновый" (медленный) сон, в котором различают собственно медленный сон и дельта-сон (около 1,5 часа), и "быстрый", или "парадоксальный" сон (в среднем 20 мин). (кроме изменений ЭЭГ) характерны для фазы "медленного" сна Снижение мышечного тонуса, брадикардия, замедление дыхания, понижение температуры тела, понижение секреторной и моторной функций желудочно-кишечного тракта, медленные движения глазных яблок.
Во время сна фазы чередуются. Волны ЭЭГ наиболее характерны для стадий "медленного" сна Альфа-ритм: частота 8 – 13 Гц, амплитуда примерно 50 мкВ; дельта-волны: частота 2 – 4 Гц, амплитуда до 150 мкВ. Волны ЭЭГ наиболее характерны для "быстрого" сна Бета-ритм: частота выше 13 Гц, амплитуда менее 25 мкВ. кроме изменения ЭЭГ) характерны для "быстрого" сна резкое снижение мышечного тонуса, быстрые движения глазных яблок, сердечная и дыхательная тахиаритмии, нарушение терморегуляции, что может сопровождаться как повышением, так и понижением температуры тела, отсутствие активности желудочно-кишечного тракта, эрекция(в фазе быстрого сна человек видит сновидения).
Цикол сна совокупность фаз "медленного" и "быстрого" сна,один цикл имеет продолжительность 80 – 100 минут.Количество циклов за ночь 4 – 5. физиологическое значение сна - отдых организма, приспособление к изменению суточной освещенности (циркадианный биоритм), переработка и запоминание новой информации.








