 2018-01-21
2018-01-21 1617
1617Секретин -стимулирует обильное сокоотделение и секрецию бикарбонатов. Секретин образуется в двенадцатиперстной кишке, а его высвобождение в кровь дуоденальными клетками происходит при переходе в нее кислого желудочного содержимого. Чем больше свободных ионов Н+ в двенадцатиперстной кишке, тем больше высвобождается секретина и тем выше объем панкреатического сока и секреция гидрокарбонатов.
Холецистокинин -усиливает секрецию поджелудочной железы, высвобождаясь в кровь из ССК-клеток слизистой оболочки 12-перстной кишки.
Секрецию поджелудочной железы усиливают также гастрин, серотонин, инсулин, бомбезин, соли желчных кислот. Тормозят выделение поджелудочного сока глюкагон, соматостатин, вазопрессин, энкефалин, кальцитонин. Вазоинтестинальный пептид может как возбуждать, так и тормозить панкреатическую секрецию.
№ 5( Физиологические основы гуморальной регуляции) Эндокринная функция надпочечников. Гормоны мозгового и коркового слоя надпочечников, их значение для функций организма. Участие гормонов надпочечников в приспособительных реакциях организма.
Надпочечники
Корковое вещество: гидрокортизон (кортизол), выработка регулируется АКТГ - Регуляция обмена углеводов, белков, жиров, глюконеогенез, катаболическое действие, липолиз, противовоспалительное действие, повышение устойчивости к инфекции.
Альдостерон - Регуляция минерального обмена и водно-солевого равновесия, увеличение активного транспорта натрия через клеточные мембраны, повышение реабсорбции натрия и воды в канальцах нефрона:задерживает в огранизме Na и Cl и усиливает выведение калия и аммония. Аналогичное влияние на клетки потовых, слюнных и кишечных желез. Участие в адаптации организма к повышенной температуре окружающей среды.
Андрогены; стероиды.
Мозговое вещество: адреналин, НА, катехоламины - Стимуляция всех видов обмена веществ, гликонеогенеза, липолиза, термогенное действие. Учащение, усиление сокращений сердца, сужение кровеносных сосудов, расширение бронхов, зрачков. Увеличение вентиляции легких, доставки кислорода к мышцам, сердцу и мозгу.
Надпочечники (корковое вещество) относятся к гипофиззависимым от передней доли гипофиза. Тропные гормоны аденогипофиза активируют выделение гормонов железам, которые воздействуют на аденогипофиз и тормозят его активность.
Мозговое вещество – гипофизнезависимые.
Участие гормонов надпочечников в приспособительных реакциях.
Важная роль в регуляции содержания глюкозы в крови в условиях гипогликемии принадлежит гормонам надпочечников. В ответ на пониженное содержание глюкозы в крови в мозговом веществе надпочечников усиливается выработка адреналина. Это является следствием первичного влияния гипогликемии на гипоталамус и гипофиз.
Роль гипоталамуса
Раздражение рецепторных клеток гипоталамуса приводит к повышению тонуса симпатико-адреналовой системы, что вызывает повышенную секреторную активность мозгового вещества надпочечников и как следствие — увеличенный выброс в кровь адреналина. Последний вместе с глюкагоном активирует фосфорилазу печени и усиливает распад печеночного гликогена. Одновременно усиливается распад гликогена мышц, поэтому после введения адреналина или избыточного его образования увеличивается концентрация глюкозы и молочной кислоты в крови.
Роль гипофиза
Стимуляция «гипогликемической кровью» гипофиза приводит к дополнительной выработке адренокортикотропного гормона. Избыточное образование адренокортикотропного гормона способствует выделению корковым веществом надпочечников глюкокортикоидов. Повышение концентрации в крови последних приводит к усилению гликонеогенеза — новообразованию глюкозы из неуглеводов, в частности из продуктов расщепления белков и жиров, что, естественно, сопровождается увеличением концентрации глюкозы в крови и содержания гликогена в печени. В мышцах и других тканях одновременно происходит усиленный распад белков, а освобождающиеся аминокислоты используются затем как исходный материал для гликонеогенеза.
№? Эндокринная функция половых желёз. Место образования половых гормонов и регуляция их продукции. Формирование и механизм половой мотивации.
Яичники
Эстрогены (эстрадиол, стероиды) - Половая дифференцировка у эмбриона, развитие половых органов, вторичных половых признаков. Обеспечение пролиферативной фазы эпителия слизистой оболочки матки. Анаболическое действие, особенно в периоде полового созревания.
Прогестерон; стероид - Подготовка слизистой оболочки матки к имплантации зародыша. Блокирует действие эстрогенов на матку, обеспечивая сохранение беременности. Торможение секреции пролактина. Катаболическое действие.
Яички
Андрогены (тестростерон, андростерон, стероиды) - Половая дифференцировка эмбриона по мужскому типу, развитие вторичных половых признаков, мужское половое поведение и агрессивность.
К гипофиззависимым относят половые железы (яички, яичники). Тропные гормоны аденогипофиза активируют выделение гормонов соответствующими железами, которые в свою очередь в определенных концентрациях воздействуют на аденогипофиз и тормозят его активность.
Женские половые органы – яичники – состоят из коркового и мозгового веществ. Гормональную функцию несет корковое вещество. С момента образования полости в фолликуле он начинает синтезировать половые гормоны. Из фолликулярной жидкости выделены три эстрогена: эстрон, эстрадиол, эстриол.
Регуляция женских половых желез осуществляется совместным действием ФСГ и ЛГ. Если ФСГ подготавливает морфологические структуры фолликула к синтезу половых гормонов, то ЛГ стимулирует образование из холестерина прегненолона — основного предшественника всех половых гормонов. В женском организме первая фаза менструального цикла, всецело связанная с развитием фолликула в яичниках, регулируется ФСГ. Этот гормон ответствен за формирование гранулезной оболочки фолликула, стимулирует гиперплазию гранулезных клеток и биосинтез ими эстрогенов. В сложном процессе разрыва созревшего фолликула доминирующую роль играет ЛГ.
Пролактин вместе с гормоном желтого тела — прогестероном — подавляет в яичниках развитие новых фолликулов. Если произошло оплодотворение, они обеспечивают имплантацию яйцеклетки и формирование плаценты. Эти гормоны ответственны за подготовку репродуктивных органов к родам, они способствуют развитию молочных желез, регулируют лактацию.
Основным источником мужских половых гормонов являются яички. Сперматогенез осуществляется в извитых канальцах. Сперматогенез регулируется в основном фолликулостимулирующим гормоном аденогипофиза.
Основными гормонами, секретируемыми яичками человека, являются тестостерон, андроапендион и дегидроэпиандростерон. Основная физиологическая роль андрогенов в мужском организме состоит в стимуляции сперматогенеза и развитии вторичных половых признаков.
Андрогены образуются не только яичками, но и надпочечниками. В репродуктивном возрасте тестостерон в мужском организме секретируется только семенниками. Андростендионобразуется и яичками, и надпочечниками.Эстрадиол образуется только яичками, а эстрон образуется из предшественников, секретируемых корковым веществом надпочечников.
В мужском организмк сперматогенез регулируется ФСГ, а биосинтез андрогенов – ЛГ.
До периода полового созревания половые гормоны вырабатываются надпочечниками. По достижении полового созревания основную роль по выработке половых гормонов берут на себя половые железы. Гормональный фон создает основу, обеспечивающую половую функцию, направленную на воспроизведение. Гормоны влияют на тканевой метаболизм, на функциональное состояние нейронов в определенных структурах мозга.
№? Функциональная система, поддерживающая оптимальный для метаболизма уровень глюкозы и крови. Характеристика её узловых механизмов.

Углеводы играют ведущую роль в энергетическом обмене организма. Деятельность практически всех без исключения органов находится в большей или меньшей зависимости от содержания углеводов в притекающей к ним крови. Местные резервы углеводов в разных тканях неодинаковы, поэтому степень зависимости скорости обменных процессов в органах и их функции определяются концентрацией глюкозы в крови.
Особенно большое значение глюкоза крови имеет для работы мышц, занимающих в количественном отношении преобладающее место в организме, и деятельности нервной системы благодаря ее ведущей, регулирующей роли в организме.
Содержание глюкозы в артериальной крови взрослого человека составляет 4,2—6,4 ммоль/л. В венозной крови содержание глюкозы обычно несколько ниже, чем в артериальной, так как часть ее переходит из крови в ткани в процессе обмена. Артериовенозная разница зависит от специфики органа и уровня его активности: в период активной работы поглощение глюкозы возрастает и артериовенозная разница увеличивается, что косвенно говорит о функциональной активности того или иного органа.
Периодические изменения уровня глюкозы в крови у человека и животных обусловлены суточными и сезонными колебаниями, приемом пищи, эмоциональным состоянием и возрастными особенностями.
Нормальный уровень глюкозы в крови, так же как и его изменения, воспринимается специальными хеморецепторами, чувствительными к изменению концентрации глюкозы крови. Глюкозорецепторы расположены в печени, сосудах, желудочно-кишечном тракте, центральной нервной системе.
Центральные глюкозорецепторы расположены в вентромедиальном отделе гипоталамуса и через рилизинг-факторы оказывают активирующее влияние на передний отдел гипофиза, а через него на деятельность таких желез внутренней секреции, как щитовидная железа, надпочечники и поджелудочная железа.
№? Функциональная система, определяющая половые функции организма. Роль биологических и социальных факторов в формировании полового поведения.
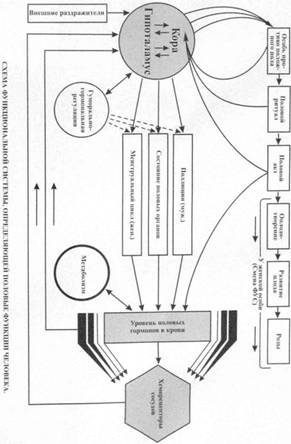
Внутренний результат - определенный уровень половых гормонов в крови. К половым гормонам относятся прежде всего мужские половые гормоны — андрогены и женские половые гормоны — эстрогены и прогестины, вырабатываемые половыми железами.
По достижении полового созревания основную роль по выработке половых гормонов берут на себя половые железы: семенники у мужских (у человека — яички) и яичники у женских особей.
Половые гормоны, кроме специфического участия в организации половых функций, оказывают на организм широкое воздействие, влияя, в частности, на процессы тканевого метаболизма, на функциональное состояние нейронов в определенных структурах мозга и т.д. Выработка половых гормонов в организме определяется не только половыми железами, но находится в тесной зависимости от внешних влияний: в женском организме в форме менструального цикла.
Гормональный фон создает как бы основу, обеспечивающую половую функцию, направленную на воспроизведение. Особенностью функциональной системы воспроизведения является то, что она строится на различных гормональных механизмах саморегуляции в женском и мужском организмах.
Внешнее звено саморегуляции - механизмы активного взаимодействия особей противоположного пола в плане достижения биологического и социального результата, осуществления полового акта, воспроизведения и продления вида.
Результатом деятельности данной системы является поддержание уровня половых гормонов, оптимального для каждого возрастного периода особей.
Гормональная и репродуктивная функции мужского и женского организма находятся под контролем сложной нейроэндокринной организации, включающей в себя гипоталамус, гипофиз, периферические железы внутренней секреции.
Первым уровнем, где реализуются гормональные эффекты, являются различные органы и ткани. Их клетки, дифференцируясь, приобретают специфические рецепторы к широкому спектру гормонов.
Среди межклеточных тканевых регуляторов ведущую роль играют простагландины. Их действие опосредуется через циклический аденозинмонофосфат.
Вторым уровнем нейроэндокринной регуляции являются периферические железы внутренней секреции. Продуцируемые ими гормоны, поступая в кровь, обладают дистантным и пролонгированным во времени действием. Воздействуя на территориально разобщенные органы, гормоны способствуют объединению их специфической деятельности. Например, молочная железа и матка обладают чрезвычайно высокой чувствительностью к половым гормонам.
Периферические эндокринные железы в свою очередь регулируются тройными гормонами гипофиза — третий уровень. Четвертый уровень составляют гипоталамические центры, которые посредством гормонов контролируют тропные функции аденогипофиза.
Пятый уровень — экстрагипоталамические влияния, непосредственно участвующие в регуляции нейросекреторных функций гипоталамических центров.
Физиология ЦНС
№ 12,1( Физиология ЦНС) Вегетативная нервная система и ее роль и регуляции различных видов деятельности организма. Особенности строения рефлекторной дуги вегетативного рефлекса. Вегетативные ганглии: локализация, классификация, механизмов передачи возбуждения в ганглиях. Вегетативные рефлексы и их виды, классификация. Адаптационно-трофическая роль вегетативных рефлексов /И.П.Павлов, Л.А.Орбели, А.Г. Гинецинский/.
Вегетативная нервная система — часть общей нервной системы, имеющая отношение преимущественно к регуляции вегетативных (растительных) функций: сокращению гладких мышц, секреции железистого аппарата, функционированию соединительной ткани внутренних органов, тонусу кровеносных и лимфатических сосудов. ВНС отчасти оказывает влияние на деятельность скелетных мышц и наружных половых органов. ВНС часто рассматривается как автономная, так как ее деятельность в слабой степени регулируется произвольно.
ВНС характеризуется:
• меньшей возбудимостью;
• меньшей функциональной лабильностью (не более 5—15 имп/с);
• меньшей скоростью проведения возбуждения;
• пролонгированным до 150 мс (за счет следовых фаз) потенциалом действия;
• временной и пространственной суммацией возбуждений в ганглиях.
Эффекторные волокна ВНС иннервируют практически все ткани и внутренние органы. Часть органов имеет «двойную» и «тройную» иннервацию: симпатическую, парасимпатическую и метасимпатическую.
В отличие от соматической нервной системы, которая построена по сегментарному принципу, вегетативная нервная система не имеет сегментарной организации. Нейроны вегетативной нервной системы располагаются в различных отделах центральной нервной системы, образуя так называемые вегетативные центры.
Вегетативная рефлекторная дуга состоит из трех звеньев, имеющих более сложное строение, чем рефлекторная дуга соматической нервной системы.
Рефлекторная дуга ВНС состоит из чувствительного (афферентного, сенсорного), ассоциативного (вставочного) и эффекторного (эфферентного) звеньев. Чувствительное (первое) звено образовано клетками спинномозговых или периферических ганглиев. Ассоциативное (второе) звено представлено преганглионарными нейронами, расположенными в боковых рогах спинного мозга, в продолговатом и в среднем мозге. Отростки вставочных преганглионарных нейронов выходят из спинного мозга в составе вентральных корешков, вступают в соматические нервные стволы и отсюда достигают узлов пограничного симпатического ствола или парасимпатических узлов в органах, где переключаются на эффекторные клетки. Эффекторное (третье) звено образовано эффекторными клетками периферических ганглиев.
Периферическая (исполнительная) часть ВНС представлена преганглионарными и ганглионарными нейронами.
Смотри №80.
Особенности строения симпатических и парасимпатических нервов. Симпатические нервы сформированы в основном преганглионарными, а парасимпатические — постганглионарными нервными волокнами. Преганглионарные волокна симпатической и парасимпатической нервной системы представлены тонкими миелиновыми волокнами типа В, постганглионарые — тонкими безмиелиновыми волокнами типа С.
К метасимпатической нервной системе относится комплекс микроганглионарных образований, расположенных в стенках внутренних органов, обладающих двигательной активностью — сердца, бронхов, мочевого пузыря, пищеварительного тракта, матки, желчного пузыря и желчных путей.
Нейроны ВНС участвуют в рефлекторных реакциях, называемых вегетативными рефлексами. Некоторые из них используются в качестве функциональных проб — глазосердечный рефлекс, дыхательно-сердечный рефлекс, ортостатическая реакция, кожный дермографизм.
Л.А.Орбели, А.Г. Гинецинский обнаружили, что раздражение постганглионарных волокон симпатического нерва восстанавливает сокращение поперечнополосатой мышцы, утомленной раздражением двигательного нерва. Этот эффект обусловлен прямым действием симпатической нервной системы на обмен веществ мышечной ткани и не связан с сосудистыми влияниями.
№ 12,11( Физиология ЦНС )Симпатический отдел вегетативной нервной системы. Функциональная организация симпатической нервной системы и ее роль врегуляции функций организма. Особенности строения рефлекторной дуги симпатической нервной системы, (расположение центров, локализации ганглиев, виды нервных волокон: медиаторы и рецепторы). Влияние парасимпатической нервной системы на функции организма.
Симпатический отдел. Тела первых нейронов симпатического отдела ВНС расположены преимущественно в задних ядрах гипоталамуса, среднем и продолговатом мозге и в передних рогах спинного мозга, начиная с 1-го грудного и кончая 3—4-м сегментом поясничного ее отдела.
Периферическая (исполнительная) часть ВНС представлена преганглионарными и ганглионарными нейронами.
Симпатический отдел. Тела преганглионарных нейронов располагаются в сером веществе боковых рогов спинного мозга, начиная от 1-го грудного сегмента до 2—3-го поясничного сегмента, и в двигательных ядрах черепных нервов. Миелиновые аксоны этих нейронов представлены медленнопроводящими возбуждение В-волокнами. Аксоны преганглионарных нейронов через синапсы взаимодействуют с нейронами, расположенными в периферических ганглиях.
Околопозвоночные ганглии расположены по обе стороны вдоль позвоночника в виде цепочек, составляющих правый и левый симпатические стволы. Продольно ганглии соединяются межузловыми соединительными ветвями, состоящими из миелиновых и безмиелиновых нервных волокон. Имеются и поперечные связи между ганглиями правой и левой стороны. Кроме этого, ганглии симпатического ствола соединяются со спинномозговыми нервами, благодаря чему они получают возможность иннервировать скелетную мускулатуру. Другая часть постганглионарных волокон направляется к внутренним органам.
Особенности строения симпатических и парасимпатических нервов. Симпатические нервы сформированы в основном преганглионарными, а парасимпатические — постганглионарными нервными волокнами. Преганглионарные волокна симпатической и парасимпатической нервной системы представлены тонкими миелиновыми волокнами типа В, постганглионарые — тонкими безмиелиновыми волокнами типа С.
Локализация ядер – заднее ядро гипоталамуса, нейроны боковых рогов грудных и поясничных сегментов спинного мозга.
Характеристика отростков – прерываются в ганглиях, преганглионарные короткие, тип В, постганглионарные длинные, тип С.
Зона иннервации – иннервирует все внутренние органы
Локализация – парабертебральные (пограничный столб), превертебральные (чревное, солнечное сплетение, брыжеечные узлы).
Медиатор – ацетилхолин, норадреналин.
Физиологические свойства симпатической нервной системы:
1.благодаря мультипликации в симпатических ганглиях распространяющееся из них возбуждение широко охватывает сразу несколько различных органов, т.е. является генерализованным. Эти влияния наиболее отчетливо прослеживаются при эмоциональных реакциях.
2.оказывает активирующие влиянияна функции иннервируемых органов: усиливает катаболические реакции, силу и частоту сокращений сердца, повышает артериальное давление, улучшает оксигенацию тканей, увеличивает содержания глюкозы в крови, скорость проведения возбуждения в скелетных мышцах и их тонус, расширяет бронхи, увеличивает объем легочной вентиляции; расширяет зрачки, увеличивает секрецию катехоламинов надпочечниками. При этом одновременно снижается тонус пищеварительного тракта, ослабляются процессы всасывания и ферментативного расщепления в кишечнике.
3. участвует в формировании целостных состояний, как агрессия, стресс, болевые реакции.
4. передача с пре- на ганглионарные нейроны осуществляется с помощью ацетилхолина, а на эффекторы — норадреналина.
5. представлена на периферии на значительном протяжении постганглионарными волокнами типа С, возбуждение на пути от ганглиев к органам распространяется медленнее, чем в парасимпатической системе.
6. Эффекты действия более продолжительны.
7. Электрические потенциалы в симпатических ганглиях характеризуются продолжительными следовыми явлениями. При стимуляции преганглионарных волокон регистрируется быстрый возбуждающий постсинаптический потенциал, который сменяется медленным тормозным постсинаптическим потенциалом.
8. начальная деполяризация обусловлена действием ацетилхолина на никотиновые рецепторы.
№ 12( Физиология ЦНС) Парасимпатический отдел вегетативной нервной системы. Структурно-функциональная организация парасимпатической нервной системы и ее роль в регуляции функций ворганизме Особенности строения рефлекторной дуги парасимпатической нервной системы (расположение центров в ЦНС, локализация ганглиев, медиаторы и рецепторы). Влияние парасимпатической нервной системы на функции различных органов.
Парасимпатический отдел Центральные нейроны расположены в передних отделах гипоталамуса, среднем и продолговатом мозге, во 2-4-м сегментах крестцового отдела спинного мозга.
Периферическая (исполнительная) часть ВНС представлена преганглионарными и ганглионарными нейронами.
Парасимпатический отдел. Тела преганглионарных нейронов расположены в среднем и продолговатом мозге среди двигательных нейронов черепных нервов: глазодвигательного, лицевого, языкоглоточного и блуждающего, а также в сером веществе боковых рогов крестцового отдела спинного мозга.
Вместе с черепными нервами преганглионарные волокна распространяются к ганглионарным нейронам, расположенным в крылонебном, подчелюстном и ушном ганглиях. Отходящие от них постганглионарные волокна иннервируют глазное яблоко, аккомодационную мышцу и сфинктер зрачка, поднижнечелюстную, подъязычную и околоушную слюнные железы.
Преганглионарные волокна блуждающего нерва распространяются к нейронам, расположенным в ганглиях органов грудной и верхней части брюшной полости.
Аксоны нейронов крестцового отдела спинного мозга через тазовые нервы адресуются к нейронам, располагающимся в ганглиях органов нижней части брюшной полости и таза, иннервируя нижнюю часть толстой кишки, прямую кишку, мочевой пузырь, нижнюю часть мочеточника и наружные половые органы.
Особенности строения симпатических и парасимпатических нервов. Симпатические нервы сформированы в основном преганглионарными, а парасимпатические — постганглионарными нервными волокнами. Преганглионарные волокна симпатической и парасимпатической нервной системы представлены тонкими миелиновыми волокнами типа В, постганглионарые — тонкими безмиелиновыми волокнами типа С.
Локализация ядер – передний гипоталамус, средний мозг, мост, продолговатый мозг, боковые рога I-V крестцовых сегментов спинного мозга.
Характеристика отростков -прерываются в ганглиях, преганглионарные длинные, тип В, постганглионарные короткие, тип С.
Зона иннервации – ограничена (нет в надпочечниках и стенках большинства сосудов)
Локализация –в иннервируемых органах (интрамурально) или рядом с ними.
Медиатор -ацетилхолин
Физиологические свойства парасимпатической нервной системы:
1. влияния направлены и локальны. Оказывает ограниченное воздействие в пределах иннервируемого органа.
2. оказывает успокаивающее, расслабляющее действие на большинство функций организма; снижается возбудимость ЦНС и миокарда, уменьшаются интенсивность метаболизма, сила и частота сердечных сокращений, кровяное давление, объем легочной вентиляции, температура тела; увеличивается секреция инсулина и как следствие снижается концентрация глюкозы в крови и увеличивается внутриклеточное депонирование ее в виде гликогена. При этом одновременно усиливаются моторная, секреторная и всасывательная функции желудочно-кишечного тракта.
3. усиливает анаболические реакции.
4.Парасимпатические влияния доминируют в формировании сна и психологического субъективного чувства удовлетворения.
5.Медиатором в преганглионарных и постганглионарных волокнах служит ацетилхолин.
6.Парасимпатическая нервная система представлена на значительном расстоянии преганглионарными волокнами типа В. Вследствие этого возбуждения от центра до органа доходят быстрее, чем по симпатическим нервам.
7.Эффекты действия парасимпатических нервов менее продолжительны.
№ 3( Физиология ЦНС) Нервный центр. Понятие об анатомическом и функциональном нервном центре. Физиологические свойства и принципы распространения возбуждения по нейронным цепям нервного центра.
№8( Физиология ЦНС )Продолговатый мозг, его нейронная организация, участие в процессах саморегуляции функции организма.
Интегративная деятельность продолговатого мозга обусловлена широким спектром афферентных потоков, поступающих от многочисленных экстеро- и интерорецепторов. Она проявляется в многочисленных реакциях, эффекторами в которых являются как скелетные и гладкие мышцы, так и железы. В продолговатом мозге находятся ядра IX и X пар черепных нервов (языкоглоточный и блуждающий нервы) и XI и XII пар черепных нервов (добавочный и подъязычный нервы).
С участием ядер черепных нервов осуществляются такие врожденные сложноорганизованные пищевые реакции, как сосание, глотание, жевание. На уровне продолговатого мозга формируются защитные реакции — чиханье, кашель, рвота, глотание, мигание, слезоотделение.
Обширны связи продолговатого мозга с хеморецепторами и барорецепторами сосудов, интерорецепторами внутренних органов и вестибулорецепторами. Влияние этих органов определяет функционирование на уровне продолговатого мозга дыхательного и сердечно-сосудистого центpoв.
В продолговатом мозге находятся ретикулярные структуры, которые дают начало ретикулоспинальному тракту, контролирующему сегментарные двигательные реакции спинного мозга. Нисходящие влияния ретикулярной формации продолговатого мозга возбуждают α- и γ- мотонейроны сгибателей и тормозят мотонейроны разгибателей, что обусловливает адекватное перераспределение тонуса мышц и амплитуды их сокращения.
№ 10( Физиология ЦНС) Ретикулярная формация: ее строение, локализация в центральной нервной системе. Центры ретикулярной формации и их участие в регуляции функций организма. Особенности восходящих и нисходящих влияний ретикулярной формации.
Морфофункциональная организация. Встволовой части мозга, куда входит и средний мозг, расположена структура, состоящая из нейронов с короткими отростками и обозначаемая как сетевидная, или ретикулярная, формация. В ней выделяют две части: каудальную, входящую в состав продолговатого мозга и моста, и ростральную, или мезэнцефальную, ретикулярную формацию. Первая часть дает начало ретикулоспинальному пути, влияющему на спинальные реакции. Мезэнпефальная ретикулярная формация дает начало восходящей активирующей системе, включающей неспецифические ядра таламуса.
Восходящие активирующие влияния. Ретикулярная формация среднего мозга осуществляет восходящие активирующие генерализованные влияния на кору большого мозга.При электрическом раздражении ретикулярной формации наблюдается переход низкочастотной высокоамплитудной электрической активности коры в низкоамплитудную высокочастотную активность. Такие же изменения электрической активности головного мозга наблюдаются при переходе организма от состояния сна к бодрствованию или при воздействии на организм самых различных раздражителей. Активное состояние ретикулярной формации всегда поддерживается непрерывным потоком афферентных импульсов, поступающих в ретикулярную формацию по коллатеральным волокнам от проекционных сенсорных проводящих путей.
Благодаря этому при воздействии раздражителей в кору большого мозга всегда поступает два потока возбуждений. Один из них направляется в кору большого мозга по проекционным сенсорным путям и достигает специфической для данного раздражителя проекционной области. Другой поток возбуждений генерализованно направляется от ретикулярной формации к коре, усиливая ее активированное состояние. Генерализованное активирующее влияние ретикулярной формации является непременным условием поддержания бодрствующего состояния мозга. Лишение коры большого мозга источника возбуждающей энергии, каковым является ретикулярная формация, приводит к переходу головного мозга в недеятельное состояние, сопровождаемое медленноволновой высокоамплитудной электрической активностью, характерной для состояния сна.
Неспецифические и специфические влияния ретикулярной формации. В связи с тем что генерализованная активация коры большого мозга возникает при любом афферентном воздействии, восходящие активирующие влияния ретикулярной формации считаются неспецифическими, не связанными со спецификой действующего раздражителя. В то же время при формировании целостных приспособительных реакций организма восходящие активирующие влияния ретикулярной формации на кору головного мозга имеют специфический характер, т.е. включены в нейрофизиологические механизмы формирования конкретной мотивации — пищевой, половой, оборонительной.
Функциональная активность ретикулярной формации обеспечивается не только обширной мультисенсорной конвергенцией возбуждений к ее нейронам и конвергенцией возбуждений, поступающих от коры большого мозга, мозжечка и других подкорковых структур, но и многочисленными гуморальными факторами, по отношению к которым ретикулярная формация обладает высокой чувствительностью. Такие гуморальные регуляторы, как адреналин и С02, являются мощными возбудителями нейронов ретикулярной формации. Нейроны ретикулярной формации содержат: норадреналин, серотонин и дофамин.
Связь с другими структурами мозга. Ретикулярная формация имеет функциональные и анатомические связи с гипоталамусом, таламусом, продолговатым мозгом и другими отделами головного мозга, поэтому все наиболее общие функции организма, такие как терморегуляция, пищевые и болевые реакции, регуляция постоянства внутренней среды организма, сон и бодрствование, находятся в функциональной зависимости от свойств ретикулярной формации ствола мозга.
№ 8( Физиология ЦНС) Средний мозг, его строение и функции. Роль среднего мозга в регуляции топических рефлексов. Децеребрационная регидность и причины её возникновения.
Средний мозг: выделяют крышу и ножки. Полостью среднего мозга является водопровод мозга. Верхней (передней) границей среднего мозга на его вентральной поверхности служат зрительные тракты и сосцевидные тела, на задней — передний край моста. На дорсальной поверхности верхняя (передняя) граница среднего мозга соответствует задним краям (поверхностям) таламусов, задняя (нижняя) — уровню выхода корешков блокового нерва.
Крыша среднего мозга расположена над водопроводом мозга. Крыша среднего мозга состоит из четырех возвышений — холмиков. Последние отделены друг от друга бороздками. Верхние холмики крыши среднего мозга (четверохолмия) и латеральные коленчатые тела выполняют функцию подкорковых зрительных центров. Нижние холмики и медиальные коленчатые тела являются подкорковыми слуховыми центрами.
Ножки мозга выходят из моста. На медиальной поверхности каждой из ножек мозга располагается продольная глазодвигательная борозда, из которой выходят корешки глазодвигательного нерва. В покрышке среднего мозга залегают ядра среднего мозга и проходят восходящие проводящие пути. Основание ножки мозга целиком состоит из белого вещества, здесь проходят нисходящие проводящие пути.
В среднем мозге расположены подкорковые центры слуха и зрения, обеспечивающие иннервацию произвольных и непроизвольных мышц глазного яблока, а также среднемозговое ядро V пары. Через средний мозг проходят восходящие (чувствительные) и нисходящие (двигательные) проводящие пути.
Основные интегративные функции среднего мозга связаны с организацией двигательных актов и анализом афферентных потоков возбуждения.
Сторожевые реакции: осуществление организмом сторожевых, или старт-реакций. Легкая степень таких реакций у человека выражается вздрагиванием при неожиданном звуке или прикосновении; при более сильных неожиданных раздражителях человек вскрикивает, а иногда даже бежит. Старт-реакции обеспечивают мгновенную мобилизацию всего организма к активной деятельности при возникновении опасности. Нейрофизиологический механизм осуществления старт-реакций связан с функционированием бугорков четверохолмия. Нейроны верхних бугорков четверохолмия обеспечивают организацию ориентировочного поведения на зрительные стимулы. Нейроны нижних бугорков четверохолмия организуют ориентировочные двигательные реакции на звук.
Тонические реакции: связаны с перераспределением тонуса различных групп мышц. К этим структурам относятся красное ядро и латеральное преддверное ядро. Тонические реакции возникают при изменении положения тела или отдельных его частей (например, головы) в пространстве. Они предотвращают нарушение равновесия тела или восстанавливают уже нарушенное равновесие.
Реакции установки тела. Совокупность тонических реакций называется реакциями установки тела. Они делятся на две группы: статические и стато-кинетические. Статические реакции возникают при изменении положения тела, не связанном с его перемещением в пространстве. При этом изменяется тонус различных групп мышц для поддержания естественной позы в случае ее изменения. Статокинетические реакции проявляются в перераспределении тонуса скелетных мышц, которое обеспечивает сохранение равновесия тела человека при угловых и линейных ускорениях активного или пассивного перемещения его в пространстве. Подобные ситуации возникают при естественных передвижениях человека — ходьбе, беге. Восприятие направления и силы ускорения, осуществляется рецепторами отолитового аппарата и полукружных каналов лабиринта внутреннего уха. Сигналы, поступающие в средний мозг от вестибулорецепторов, вызывают вращательные реакции глаз, головы, конечностей и туловища.
Перерезка у животного ствола мозга между передними и задними буграми четверохолмия (операция перерезки ствола мозга называется децеребрацией) вызывает состояние скелетной мускулатуры, которое называется децеребрационной ригидностью. Это состояние характеризуется резким повышением тонуса разгибательной мускулатуры. Конечности сильно вытянуты, голова запрокинута, спина выгнута.
№? Промежуточный мозг. Гипоталамус, его функциональная организация. Роль гипоталамуса в регуляции гомеопатических, вегетативных и эндокринных функций организма, в формировании врождённых реакции, в механизмах эмоции и мотивации.
Границами промежуточного мозга на основании головного мозга являются сзади — передний край заднего продырявленного вещества и зрительные тракты, спереди — передняя поверхность зрительного перекреста. На дорсальной поверхности задней границей является борозда, отделяющая верхние холмики среднего мозга от заднего края таламусов. Переднебоковая граница разделяет с дорсальной стороны промежуточный мозг и конечный.
Промежуточный мозг включает следующие отделы: таламическую область (область зрительных бугров, зрительный мозг), гипоталамус, объединяющий вентральные отделы промежуточного мозга; III желудочек.
Структуры ЦНС, обеспечивающие регуляцию деятельности внутренних органов, поддерживающие постоянство внутренней среды организма и формирующие мотивационные состояния организма, объединяются понятием «висцеральный мозг». Он включает гипоталамус и лимбические образования ЦНС.
Гипоталамус является структурой ЦНС, осуществляющей сложную интеграцию и приспособление функций различных внутренних органов к целостной деятельности организма. Гипоталамус объединяет и связывает в единое целое механизмы гуморальной и нервной регуляции. Под контролем гипоталамуса находятся такие железы внутренней секреции, как гипофиз, щитовидная, половые железы, надпочечники и др. Регуляция тропных функций гипофиза осуществляется путем выделения гипоталамическими нейронами гормонов, поступающих в гипофиз в основном через портальную систему сосудов. Выделение тропных гормонов гипофиза приводит к изменению функций эндокринных желез, секрет которых попадает в кровь и в свою очередь может действовать на гипоталамус (обратная связь).
Передняя область гипоталамуса принимает непосредственное участие в регуляции выделения гонадотропинов и оказывает стимулирующее влияние на половое развитие организма. Гормоны нейрогипофиза являются продуктом секреции супраоптического ядра гипоталамуса (например, вазопрессин или антидиуретический гормон).
Регуляция вегетативных функций. Под контролем гипоталамических центров находятся такие интегративные функции организма, как поддержание постоянства температуры тела, углеводный, жировой и водный обмены организма, регуляция давления крови, регуляция половых функций и функций желудочно-кишечного тракта и др.
В зависимости от выполняемых функций в гипоталамусе выделяют две зоны. Первой зоной является динамогенная, занимающая среднюю и заднюю части гипоталамуса. При ее возбуждении наблюдаются расширение зрачка, повышение кровяного давления, активация дыхания, повышение двигательной возбудимости, т.е. проявления симпатических влияний вегетативной нервной системы. Второй зоной является трофогенная, находящаяся в преоптической области гипоталамуса. Возбуждение ее проявляется в сужении зрачка, снижении кровяного давления, урежении дыхания, рвоте, дефекации, мочеиспускании, слюноотделении, т.е. симптомах, характерных для влияний парасимпатической нервной системы.
В гипоталамусе располагаются центры голода, насыщения, жажды и др. Получая афферентные потоки возбуждений от интерорецепторов (осморе-цепторов, хеморецепторов, терморецепторов и т.д.) и интегрируя их с гуморальными влияниями на нервные клетки гипоталамуса, эти центры формируют соответствующие мотивационные состояния организма.
Гипоталамус относится также к гипногенным структурам ЦНС, которые в функциональном взаимодействии обеспечивают смену сна и бодрствования.
№ 10( Физиология ЦНС )Мозжечок. Морфологическая и функциональная организация коры и ядер мозжечка. Участие мозжечка в регуляции познотонических рефлексов и висцеральных функций организма.
Мозжечок располагается дорсальнее от моста и от верхней дорсальной части продолговатого мозга. Он лежит в задней черепной ямке.
Интрегративные фкнкции мозжечка связаны с организацией двигательных актов и регуляцией вегетативных функций. При осуществлении двигательного акта перемещающиеся части тела испытывают влияние инерционных сил, что нарушает плавность и точность выполняемого движения. Коррекция движения осуществляется структурами мозжечка. В промежуточную часть мозжечка по коллатералям кортико-спинального тракта поступает информация о планируемом движении, а также афферентация от соматосенсорной системы. В результате формируется возбуждения к красному ядру и стволовым двигательным центрам, обеспечивающие взаимную координацию движений.
Особенно большое значение мозжечок имеет для построения быстрых баллистических целенаправленных движений Коррекция формируется в полушариях мозжечка и его зубчатом ядре на основе импульсации, поступающей от всех областей коры большого мозга, и фиксируется в мозжечке. Связь мозжечка с высшими вегетативными центрами и с некоторыми железами внутренней секреции обеспечивает его участие в регуляции вегетативных функций. Мозжечок оказывает стабилизирующее влияние на деятельность пищеварительного тракта, дыхание, деятельность сердца и тонус сосудов, терморегуляцию, обмен веществ.
Центральное место среди структур экстрапирамидной системы занимают базальные ядра. При их участии осуществляется синергизм всех элементов таких сложных двигательных актов, как ходьба, бег, лазанье; достигаются плавность движений и установка исходной позы для их осуществления.
Функции полосатого тела и бледного шара. Среди структурных образований экстрапирамидной системы полосатое тело считается высшим подкорковым регуляторно-координационным центром организации движений. Полосатое тело и бледный шар, влияя на нейроны спинного мозга через структуры среднего и продолговатого мозга, координируют тонус и фазовую двигательную активность мышц. Бледный шар оказывает тормозяшее воздействие на ядра среднего мозга.
В отличие от полосатого тела (стриатума) неостриатумвключает хвостатое ядро и скорлупу. Эти образования вызывают торможение моторного компонента условных и безусловных реакций организма.
Особенности морфофункциональной организации. Базальные ядра не имеют прямых выходов к мотонейронам спинного мозга, а опосредуют свои влияния на них через ретикулоспинальный тракт, являющийся как бы общим конечным путем. Эти влияния адресуются к γ-мотонейронам спинного мозга, которые регулируют поток проприоцептивных афферентных импульсов, поступающих в спинной мозг от мышечных веретен. Эти афферентные импульсы влияют на возбудимость а-мотонейронов, активность которых определяет рабочее состояние скелетных мышц.
Хвостатое ядро, скорлупа и бледный шар участвуют не только в регуляции моторной деятельности, но и в анализе афферентных потоков, в регуляции ряда вегетативных функций, в осуществлении сложных форм врожденного поведения, в механизмах кратковременной памяти, а также в регуляции цикла бодрствование—сон. На нейронах базальных ядер происходит взаимодействие афферентных потоков, поступающих практически от всех сенсорных структур, от многих областей коры головного мозга, от таламических, ретикулярных, лимбических и других структур мозга.
Таким образом, широкие афферентные и эфферентные связи базальных ядер между собой, их двусторонние связи с корой большого мозга, особенно с ее моторными зонами, а также связи со структурами промежуточного, среднего и продолговатого мозга обеспечивают широкое взаимодействие возбуждений на нейронах, что является основой высшей интеграции и контроля поведенческих актов.
№ 11( Физиология ЦНС)? Кора больших полушарий. Методы исследования. Современные представления о локализации функций в коре головного мозга. Особенности деятельности левого и правого полушарии коры головного мозга Роль коры в произвольной регуляции двигательной активности.
Морфофункциональная организация. Вкоре большого мозга различают пять долей: лобную, теменную, затылочную, височную и островковую, каждая из которых имеет проекционные и ассоциативные области. К проекционным областям коры импульсы возбуждения поступают преимущественно от специфических сенсорных и двигательных ядер таламуса. Основным источником возбуждений, поступающим к ассоциативным областям коры мозга, являются другие проекционные и ассоциативные корковые зоны.
Проекционные и ассоциативные области коры мозга получают сигналы от неспецифических ядер таламуса. Эти неспецифические влияния определяют уровень активного состояния коры большого мозга.
Представления о функциональной организации различных областей коры большого мозга получены при микроэлектродной регистрации биоэлектрической активности отдельных нейронов.
Особенности проекционных зон. Общим является наличие большого количества специфических нейронов, которые дают реакции на раздражители строго определенной сенсорной модальности. Среди специфических нейронов выделены проекционные нейроны, имеющие однозначную связь с проекционным рецептивным периферическим полем, и непроекционные нейроны, возбуждающиеся с различных рецептивных полей одной модальности. Например, некоторые нейроны зрительной области коры большого мозга реагируют на звуковые стимулы. Неспецифические нейроны, как правило, больше находятся в ассоциативных областях коры головного мозга.
Одним из общих механизмов функционирования нейронов различных областей коры мозга является механизм конвергенции возбуждений к отдельным нервным клеткам.
Мультисенсорная конвергенция. В коре большого мозга особенно многочисленны эффекты мультисенсорной конвергенции, поскольку в кору головного мозга приходят все афферентные потоки возбуждений. Чаше всего эффект мультисенсорной конвергенции дают неспецифические нейроны, что проявляется в реакции отдельных нервных клеток на несколько предъявляемых раздражителей (звуковой, световой, соматосенсорный и др.).
Сенсорно-биологическая конвергенция проявляется в схождении к отдельным нейронам коры большого мозга мотивационных возбуждений, связанных с различными биологическими состояниями организма (боль, голод и др.).
Мультибиологическая конвергенция. Мотивационные состояния организма возникают на основе генерализованных восходящих активирующих влияний подкорковых образований на кору большого мозга. Эти восходящие влияния, формируемые подкорковыми структурами — гипоталамусом, ретикулярной формацией, лимбическими образованиями.
Эфферентно-афферентная конвергенция -является одним из механизмов формирования в ЦНС аппарата предвидения результатов поведенческого акта — акцептора результатов действия.
Два полушария большого мозга объединяются мозолистым телом, волокна которого связывают идентичные пункты коры большого мозга и обеспечивают единство ее функционирования. У большинства людей доминирующим является левое полушарие, которое обеспечивает функцию речи, контроль за действием правой руки, вербальное, логическое мышление. Такой человек тяготеет к теории, имеет большой запас слов, ему присущи целеустремленность, повышенная двигательная активность, способность предвидеть события.
Правое полушарие головного мозга специализировано для восприятия формы и пространства и участвует в интуитивном мышлении. Доминирование правого полушария проявляется у человека в конкретных видах деятельности, в способности тонко чувствовать и переживать.
Анализаторы
№ 1( Анализаторы) Физиология анализаторов. Понятие о сенсорных системах. Строение анализаторов, свойства периферического, проводникового и центрального отделов анализатора. Роль анализаторов в деятельности функциональных систем организма.
Анализатор – совокупность возбудимых структур центральной и периферической нервной системы, осуществляющих восприятие и анализ воздействий окружающей среды и воздействий, исходящих от самого организма.
Понятие о сенсорных системах:
Системы организма, воспринимающие раздражения из окружающей среды с помощью экстерорецепторов. К ним относятся фоторецепторы, слуховые, тактильные, температурные и хеморецепторы, расположенные на поверхностях тела и в начальных отделах пищеварительного тракта и дыхательных путей.
Все структуры, входящие в состав анализаторов, относятся к афферентным, т.е. проводящим возбуждения от периферии в ЦНС. Классические представления Павлова об анализаторе включают в его состав три части: периферический отдел, проводниковый отдел и центральный конец.
Периферический отдел анализаторов включает, как правило, рецепторы, хотя в некоторых анализаторах, например зрительном, в этот отдел могут быть включены и первичные афферентные нейроны. Периферический отдел анализатора является составной частью любого органа чувств, который, помимо рецепторов, включает специальные вспомогательные образования для наилучшего восприятия действующего раздражителя. Например, глаз как орган зрения, помимо сетчатки (фоторецепторы), включает глазное яблоко, его мышцы, веки и др.
Проводниковый отдел анализаторов включает не только нервные волокна, непосредственно отходящие от рецепторов, но и все афферентные нейроны, обеспечивающие первичный анализ и передачу возбуждений в центральный отдел анализатора. Возникающие в рецепторах импульсы возбуждения распространяются по проводящим путям в виде электрических потенциалов. Во всех нервных волокнах потенциалы являются однотипными по внешнему виду, но в потоке импульсов возбуждения в их своеобразном рисунке — паттерне — закодирована специфическая информация о параметрах действующего раздражителя. Анализ этой информации начинается как на уровне первичных афферентных нервных клеток, так и в последующих спинальных, стволовых и подкорковых ядрах.
Центральный отдел анализаторов. Различные проводящие афферентные пути через возбуждение соответствующих подкорковых структур в конечном счете приносят импульсы возбуждения в соответствующие области коры большого мозга, которые считаются высшим центральным конечным звеном любого анализатора. Вместе со специфическим афферентным возбуждением в кору поступает и неспецифическое восходящее возбуждение, которое формируется на уровне подкорковых активирующих структур мозга — ретикулярной формации, гипоталамуса и др.
Передача импульсов от рецепторов по проводящим путям к коре большого мозга осуществляется по цепям нейронов в различных ядрах, расположенных на разных уровнях ЦНС. За счет конвергенции и дивергенции возбуждений в нейронных цепях в этих нервных центрах осуществляются передача и обработка информации.
Роль анализаторов в деятельности функциональных систем:
Физиологические особенности каждого анализатора в отдельности определяются его специфическими структурами передачи возбуждений от рецепторов в ЦНС, участием в системных процессах целого организма. Адекватное поведение живых организмов в окружающей среде не является пассивным отражением воздействующих раздражителей. В большей степени организм настойчиво ищет потребные раздражители и активно к ним стремится, избирательно настраивая по отношению к ним свои анализаторы.
Активное стремление субъектов к раздражителям внешней среды определяется прежде всего их исходными доминирующими потребностями и пропускной способностью к передаче информации соответствующего анализатора. У человека наибольшей пропускной способностью обладает зрительный анализатор, который в единицу времени передает в ЦНСболее 70 % информации; 25—28 % информации доставляет в ЦНС слуховой анализатор и 2—5 % информации — остальные анализаторы.
№? Рецепторы. Классификация рецепторов. Основные свойства рецепторов. Закон Вебера-Фехтенера и его анализ.
Рецепторы участвуют впроцессе восприятия и трансформации механической, термической, электромагнитной и химической энергии в нервный сигнал или сложную последовательность мембранных и цитоплазматических процессов.
Существуют различные классификации рецепторов, основанные на их физиологических характеристиках.
Психофизиологическое состояние, связанное с модальностью ощущения, в соответствии с которым выделяют зрительные, слуховые, осязательные, обонятельные, вкусовые, холодовые, тепловые, болевые рецепторы.
Локализация. Большинство сенсорных рецепторов воспринимает раздражения из окружающей среды, т.е. являются внешними, или экстерорецепторами. К ним относятся фоторецепторы, слуховые, тактильные, температурные и хеморецепторы, расположенные на поверхности тела и в начальных отделах пищеварительного тракта и дыхательных путей.
Информация о состоянии внутренней среды организма воспринимается интерорецепторами внутренних органов, сосудов, опорно-двигательного аппарата — мышц, сухожилий, костей, суставов.
Структура рецепторов. Рецепторы могут быть представлены свободными нервными окончаниями; окончаниями, покрытыми особой капсулой (инкапсулированные; иметь вид палочек, колбочек, ветвей, волосков. Некоторые рецепторы объединяются в сложно организованные множества — сетчатку глаза, кортиев орган внутреннего уха и др. В результате рецепции действующего на организм раздражения и поступающей в мозг на ее основе сигнализации формируется субъективно переживаемое ощущение, являющееся источником познания внешнего мира.
Специализация рецепторов. Характерным свойством рецепторов является их высокая генетически детерминированная специализация к восприятию адекватного раздражителя. В соответствии с природой или характером раздражения их делят на:
• тактильные рецепторы кожи;
• слуховые, вестибулярные и гравитационные рецепторы внутреннего уха;
• рецепторы опорно-двигательного аппарата (растяжения, суставные, мышц);
• барорецепторы сердца и сосудов;
• хеморецепторы обоняния, вкуса, кровеносных сосудов и тканей;
• фоторецепторы сетчатки — нервные элементы, возбуждаемые электромагнитными волнами дающие ощущения ахроматического — черно-белого (палочки) и хроматического — цветового (колбочки) видения;
• терморецепторы кожи, внутренних органов и ЦНС, реагирующие на изменения температуры окружающей среды и внутренней среды организма.
Кроме этого, выделяют рецепторы вибрации, рецепторы волосяных фолликулов, ганглиев.
Модальность. Некоторые (мономодальные) рецепторы приспособлены для восприятия лишь одного вида раздражения, например вкусовые рецепторы сладкого; другие (полимодальные) — для восприятия нескольких видов раздражителей, например ноцицепторы кожи, участвующие в формировании болевого ощущения при любом механическом, химическом, температурном повреждающем воздействии.
Дистантные - воспринимают информацию от источника, расположенного на некотором расстоянии от них (зрительные, слуховые)
Контактные — при непосредственном соприкосновении с раздражителем (тактильные).
Чувствительность. Большинство рецепторов обладает высокой чувствительностью по отношению к адекватным раздражителям
Низкопороговые — наиболее чувствительные рецепторы — расположены в коже (тактильные, или осязательные, волоски), в сетчатке глаза (палочки), в обонятельных луковицах.
К высокопороговым — наименее чувствительным — относятся рецепторы сетчатки (колбочки), ответственные за хроматическое (цветовое) зрение, и ноцицепторы кожи, возбуждающиеся при механическом воздействии повреждающей интенсивности.
Адаптация — изменение порога чувствительности рецептора при постоянном действии на него раздражителя.
Закон Вебера-Фехтенера и его анализ:
Для всех органов чувств человека ощущение пропорционально логарифму раздражителя, выраженному в единицах порога ощущения.
Возрастание силы раздражения в геометрической прогрессии стоит в соответствии росту ощущения в арифметической прогрессии. Эта формула измерения ощущений была выведена на основе исследований Вебера, в которых было показано постоянство относительной величины приращения раздражителя, вызывающего ощущение едва заметного различия. При этом был введен собственный постулат о том, что едва заметный прирост ощущения является величиной постоянной и может быть использован в качестве единицы измерения ощущения.
№ 2( Анализаторы) Зрительный анализатор. Строение вспомогательного аппарата, оптической системы и рецепторного аппарата зрительного анализатора. Фотохимические и электрические явления в сетчатке. Бинокулярное зрение, его физиологический механизм и значение. Цветное зрение. Методы изучения зрительного анализатора.
Светопреломляющие структуры глаза: роговица, радужная оболочка, хрусталик, камерная влага и стекловидное тело - обеспечивают формирование на сетчатке реального, уменьшенного и перевернутого изображения объекта внешнего мира. Радужная оболочка образует зрачок. Светопреломляющая способность хрусталика и диаметр зрачка изменяются при сокращении гладких мышц глаза. Зрачковая реакция на свет является механизмом снижения количества света, падающего на сетчатку при сильном освещении (сужение зрачка), или повышения количества света при слабом освещении за счет увеличения ширины зрачка. Физиологические механизмы опознания зрительных объектов начинаются с первичной обработки зрительной информации в сетчатке глаза, которая является периферической рецепторной структурой зрительного анализатора. Сетчатка расположена на внутренней поверхности задней сферы глазного яблока и состоит из клеток пигментного эпителия, фоторецепторов и четырех слоев, образованных различными нервными клетками.
Фоторецепторы сетчатки: основными зрительными рецепторами, расположенными в сетчатке, являются палочки и колбочки. У человека рецепторный слой сетчатки состоит из 120 млн палочек и 6 млн колбочек. Колбочки воспринимают цвета и функционируют в условиях яркой освещенности объектов, в то время как палочки воспринимают световые потоки в условиях сумерек.
Фоторецепторы сетчатки содержат светочувствительные пигменты, которые обесцвечиваются при действии света. В палочках содержится пигмент родопсин, в колбочках — йодопсин. Процесс преобразования энергии в фоторецепторе начинается с поглощения фотона молекулой пигмента. Конформационное изменение молекул пигмента активирует ионы Са2+, которые посредством диффузии достигают натриевых каналов, вследствие чего проводимость для Na+ снижается. В результате снижения натриевой проводимости возникает увеличение электроотрицательности внутри фоторецепторной клетки по отношению к внеклеточному пространству.
Сетчатка представляет собой довольно сложную нейронную сеть с горизонтальными и вертикальными связями между фоторецепторами и клетками. Биполярные клетки сетчатки передают сигналы от фоторецепторов в слой ганглиозных клеток и к амакриновым клеткам (вертикальная связь). Горизонтальные и амакриновые клетки участвуют в горизонтальной передаче сигналов между соседними фоторецепторами и ганглиозными клетками.
Зрительные пути: Аксоны ганглиозных клеток дают начало зрительному нерву. Правый и левый зрительные нервы сливаются у основания черепа, образуя перекрест, где нервные волокна, идущие от внутренних половин обеих сетчаток, пересекаются и переходят на противоположную сторону. Волокна, идущие от наружных половин каждой сетчатки объединяются вместе с перекрещенным пучком аксонов из контралатерального зрительного нерва, образуя зрительный тракт. Зрительный тракт заканчивается в первичных центрах зрительного анализатора, к которым относятся латеральные коленчатые тела, верхние бугорки четверохолмия и претектальная область ствола мозга.
Латеральные коленчатые тела являются первой структурой ЦНС, где происходит переключение импульсов возбуждения на пути между сетчаткой и корой большого мозга. Нейроны сетчатки и латерального коленчатого тела производят анализ зрительных стимулов, оценивая их цветовые характеристики, пространственный контраст и среднюю освещенность в различных участках поля зрения. В латеральных коленчатых телах начинается бинокулярное взаимодействие от сетчатки правого и левого глаза.
Верхние бугорки четверохолмия. Нервные клетки реагируют на движущиеся световые стимулы, включены в механизмы управления целенаправленным движением глаз.
Бинокулярное зрение -механизм регуляции одновременного движения правого и левого глазных яблок, который управляются нейронами, находящимися как в подкорковых структурах, так и в коре большого мозга. Центры бинокулярного зрения находятся в области ретикулярной формации среднего мозга, в верхних бугорках четверохолмия. Ретикулярная формация среднего мозга является интегрирующим центром, получающим информацию по афферентным путям не только от верхних бугорков четверохолмия, но и от фоторецепторов сетчатки. Ядра глазодвигательных нервов находятся также под влиянием мозжечка. В мозжечке вестибулярные и зрительные сигналы интегрируются с сигналами, отражающими положение головы и глаз.
Цветное зрение: восприятие глазом того или иного тона зависит от длины волны излучения: длинноволновые – красный и оранжевый; средневолновые – желтый и зеленый; коротковолновые – голубой, синий, фиолетовый. За пределами хроматической части спектра располагается невидимое невооруженным глазом ультрафиолетовое излучение. В соответствии с трехкомпонентной теорией цветового зрения нормальное ощущение цвета называется нормальной трихромазией.
Методы изучения зрительного анализатора:
1. определение остроты зрения;
2. исследование периферического (черно – белого) зрения
3. периметрия (определение поля зрения)
4. исследование бинокулярного зрения.
№ 3( Анализаторы) Слуховой анализатор. Особенности рецепторного проводникового и коркового отделов анализатора. Механизм передачи звуковых колебаний. Методы исследования слухового анализатора.
Максимальная чувствительность слуха человека лежит в области частот от 1000 до 4000 Гц.
В кортиевом органе различают внутренние и наружные волосковые клетки. Фонорецепторы кортиева органа являются вторично чувствующими рецепторами. Афферентные биполярные слуховые нейроны находятся в спиральном ганглии. От каждой клетки спирального ганглия один отросток идет на периферию к волосковым клеткам кортиева органа, а другой в составе слухового нерва направляется в ЦНС.
Звук вызывает колебания эндолимфы улиткового протока попеременно в сторону вестибулярной и в сторону барабанной лестницы. Результатом такого движения является смещение основной и покровной мембраны кортиева органа относительно друг друга. Сгибание цилий является для волосковых клеток адекватным стимулом. При этом в волосковых клетках возникает рецепторный потенциал, который вызывает высвобождение медиатора. Медиатор действует возбуждающим образом на постсинаптическую мембрану афферентного волокна биполярного нейрона спирального ганглия, что в конечном счете приводит к возникновению потенциалов действия в волокнах слухового нерва.
Отдельные участки улитки воспринимают определенные звуковые частоты. Каждое нервное волокно оптимально возбуждается звуком определенной частоты. У основания кортиева органа расположены рецепторные клетки, воспринимающие низкие звуки; у вершины улитки — рецепторы, воспринимающие высокие звуки.
Слуховые пути: п ервичные афферентные волокна распространяются сначала к вентральной и дорсальной частям кохлеарного ядра. От вентральной части вентральный тракт направляется к ипси- и контралатеральным оливарным комплексам. Таким образом, нервные клетки в каждом оливарном комплексе получают возбуждения от рецепторов правого и левого уха, что обеспечивает сравнительную оценку акустической информации. Дорсальное кохлеарное ядро служит началом дорсального слухового тракта, волокна которого переходят на противоположную сторону и там образуют синапсы с нейронами ядра латеральног

