 2014-01-25
2014-01-25 1019
1019Особенности метаболизма нервной ткани
Содержание основных минеральных компонентов в головном мозге и плазме крови
Минеральные вещества
Nа, К, Сu, Fе, Са, Мg и Мn распределены в головном мозге относительно равномерно между серым и белым веществом. Содержание фосфора в белом веществе выше, чем в сером.
| Компонент | Мозговая ткань, ммоль/кг | Плазма крови, ммоль/л |
| Na+ К+ Са2+ Сl НСОз | 2,5 101 |
Концентрация ионов Nа, К, а также Cl в мозге резко отличается от их концентрации в жидкостях тела. Соотношение неорганических анионов и катионов в мозговой ткани свидетельствует о дефиците анионов. Для покрытия дефицита анионов потребовалось бы в 2 раза больше белков, чем их имеется в ткани мозга. Принято считать, что остающийся дефицит анионов покрывается за счет липидов. Вполне возможно, что участие липидов в ионном балансе — одна из функций липидов в деятельности головного мозга.
Интенсивность обновления богатых энергией фосфорных соединений в головном мозгу очень велика, поэтому содержание АТФ и креатинфосфата в мозговой ткани очень постоянно. Полное прекращение доступа кислорода (даже на 10-15 сек при нарушении кровоснабжения мозга) повреждает энергетику нервных клеток. Это выражается обморочными состояниями.
При нормальных условиях энергетические потребности зрелого мозга обеспечиваются почти полностью за счет аэробного гликолиза.
У человека масса мозга составляет 2 % от массы тела, но на долю мозга (в состоянии покоя) приходится ≈20 % потребления О2 и ≈60 % глюкозы. Зависимость головного мозга от глюкозы означает, что резкое падение уровня глюкозы в крови, например, в случае передозировки инсулина у диабетиков, может стать опасным для жизни. Поступление глюкозы в нейроны обеспечивает мембраный белок-переносчик GLUT-3 с низкой величиной Кт (1-2 ммоль/л). Мозг эффективно поглощает глюкозу из крови и окисляет со скоростью 75 мг за 1 мин. Во всём головном мозге содержится около 750 мг глюкозы, чего хватает на 10 мин жизни, поскольку мозгу нужен постоянный приток глюкозы. Окисление глюкозы в ткани мозга до СO2 и Н2O на 90 % происходит при участии цикла Кребса и дыхательной цепи, а также путем гликолиза.
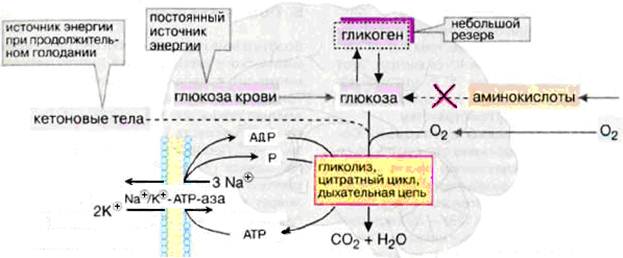
Рис. Энергетический обмен в головном мозге
При кислородном голодании головной мозг может получать энергию за счет процессов гликолиза очень недолго, так как содержание глюкозы в нем невелико. Запасы гликогена в клетках головного мозга незначительны. Жирные кислоты, которые в плазме крови транспортируются в виде комплекса с альбумином, не достигают клеток головного мозга из-за гематоэнцефалического барьера. Аминокислоты не могут служить источником энергии для синтеза АТФ, поскольку в нейронах отсутствует глюконеогенез. Только при продолжительном голодании клетки начинают использовать дополнительный источник энергии – кетоновые тела. В пренатальный и неонатальный периоды (т.е. перед рождением и в ранние сроки после рождения), а также у голодающих (независимо от возраста) на долю окисляемого ацетоацетата может приходиться ¼ потребляемого кислорода.
В связи с тем, что аэробные превращения глюкозы – главный источник энергии, важное значение приобретает реакция окислительного декарбоксилирования пирувата. Считают, что полиневриты и другие неврологические признаки недостаточности витамина В1 (значит и дефицита кофермента ТДФ) – следствие неадекватного превращения пирувата в ацетил-КоА. В результате пируват накапливается, а его высокие концентрации токсичны для нервной системы.
В клетках ЦНС до 40 % АТФ потребляет транспортная Na+/К+-АТФаза клеточных мембран. Активный транспорт Na+ и К+ компенсирует постоянный поток ионов через ионные каналы, необходимый для создания электрических потенциалов. Кроме того, АТФ используется для биосинтезов.
СЕМЕЙСТВО БЕЛКОВ: Натрий-водородный противопереносчик (Na+/H+-АТФаза мембран, белок NHE) регулятор внутриклеточного рН, участвует:
1) в обмене катионов между клетками и внеклеточной жидкостью и
2) в почечных процессах ацидогенеза.
n NНЕ5 — регулятор рН и объема клеток в ЦНС: участие в механизмах защиты от набухания и отёка мозга
n NНЕ4 — клетки слизистой желудка: участие в секреции HCI.
n NHE3 — апикальные отделы плазматических мембран эпителия кишечника и почек.
n NНЕ2 — канальцы почек и слизистой кишечника.
n NHЕ1 — на всех клетках, кодируется геном SLC9A1. Именно дефект этого гена и данного белка ведет к развитию эссенциальной гипертензии. С ним связано развитие всех проявлений метаболического Х-синдрома (гипертензии, диабета, ожирения, гиперурикемии, атеросклероза).








