 2020-01-14
2020-01-14 1134
1134Под понятием "вектор" понимается молекула нуклеиновой кислоты, способная после введения в клетку к автономному существованию за счет наличия в ней сигналов репликации и транскрипции.
Векторные молекулы должны обладать следующими свойствами:
1) способностью автономно реплицироваться в клстке-реципиенте, то есть быть самостоятельным репликоном;
2) содержать один или несколько маркерных генов, благодаря экспрессии которых у клетки-реципиента появляются новые признаки, позволяющие отличить трансформированные клетки от исходных;
3) содержать по одному или, самое большее, по два участка (сайта) для различных рестриктаз в разных районах (в том числе в составе маркерных генов), но не в области, ответственной за их репликацию.
В зависимости от целей эксперимента векторы можно условно разделить на две группы: 1) используемые для клонирования и амплификации нужного гена; 2) специализированные, применяемые для экспрессии встроенных чужеродных генов. Вторая группа векторов объединяет векторы, призванные обеспечить синтез белковых продуктов клонированных генов. Векторы для экспрессии содержат последовательности ДНК, которые необходимы для транскрипции клонированных копий генов и трансляции их мРНК в штаммах клеток.
В качестве прокариотических векторов используются плазмиды, бактериофаги; в качестве эукариотических векторов применяют вирусы животных и растений, векторы на основе 2 мкм дрожжей и митохондрий и ряд искусственно сконструированных векторов, способных реплицироваться как в бактериальных, так и в эукариотических клетках (челночные векторы).
Плазмиды - это внехромосомные генетические элементы про- и эукариот, которые автономно реплицируются в клетках. Большинство плазмидных векторов получено на основе природных плазмид ColE1, pMB1 и p15A.
Бактериальные плазмиды делят на два класса. Одни плазмиды (например, хорошо изученный фактор F, определяющий пол у E.coli) сами способны переходить из клетки в клетку, другие такой способностью не обладают. По ряду причин, и прежде всего для предотвращения неконтролируемого распространения потенциально опасного генетического материала, подавляющее большинство бактериальных плазмидных векторов создано на базе плазмид второго класса. Многие природные плазмиды уже содержат гены, определяющие устойчивость клеток к антибиотикам (продукты этих генов - ферменты, модифицирующие или расщепляющие антибиотические вещества). Кроме того, в эти плазмиды при конструировании векторов вводятся дополнительные гены, определяющие устойчивость к другим антибиотикам.
На рис. 1 показан один из самых распространенных плазмидных векторов E.coli - pBR322. Он сконструирован на базе детально изученной плазмиды E.coli - колициногенного фактора ColE1 - и содержит ориджин репликации этой плазмиды. Особенность плазмиды ColE1 (и pBR322 соответственно) состоит в том, что в присутствии ингибитора синтеза белка антибиотика хлорамфеникола (опосредованно ингибирующего репликацию хозяйской хромосомы) ее число в E.coli возрастает от 20-50 до 1000 молекул на клетку, что позволяет получать большие количества клонируемого гена. При конструировании вектора pBR322 из исходных плазмид был делегирован целый ряд "лишних" сайтов для рестриктаз.
В настоящее время наряду с множеством удобных векторных систем для E.coli сконструированы плазмидные векторы для ряда других грамотрицательных бактерий (в том числе таких промышленно важных, как Pseudomonas, Rhizobium и Azotobacter), грамположительных бактерий (Bacillus), низших грибов (дрожжи) и растений.
Плазмидные векторы удобны для клонирования относительно небольших фрагментов (до 10 тыс. пар оснований) геномов небольших размеров. Если же требуется получить клонотеку (или библиотеку) генов высших растений и животных, общая длина генома которых достигает огромных размеров, то обычные плазмидные векторы для этих целей непригодны. Проблему создания библиотек генов для высших эукариот удалось решить с использованием в качестве клонирующих векторов производных бактериофага l.
Среди фаговых векторов наиболее удобные системы были созданы на базе геномов бактериофагов l и М13 E.coli. ДНК этих фагов содержит протяженные области, которые можно делегировать или заменить на чужеродную ДНК, не затрагивая их способности реплицироваться в клетках E.coli. При конструировании семейства векторов на базе ДНК l фага из нее сначала (путем делений коротких участков ДНК) были удалены многие сайты рестрикции из области, не существенной для репликации ДНК, и оставлены такие сайты в области, предназначенной для встраивания чужеродной ДНК. В эту же область часто встраивают маркерные гены, позволяющие отличить рекомбинантную ДНК от исходного вектора. Такие векторы широко используются для получения "библиотек генов". Размеры замещаемого фрагмента фаговой ДНК и соответственно встраиваемого участка чужеродной ДНК ограничены 15-17 тыс. нуклеотидных остатков, так как рекомбинантный фаго - вый геном, который на 10% больше или на 75% меньше генома дикого l фага, уже не может быть упакован в фаговые частицы.
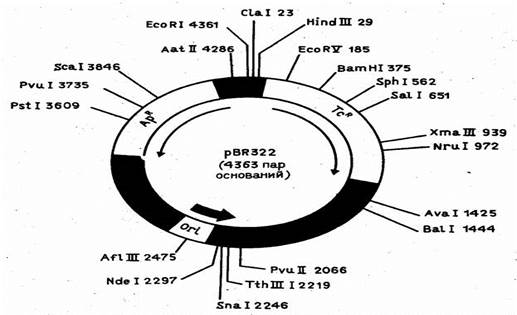
Рисунок 1. Детальная рестрикционная карта плазмиды pBR322.
Таких ограничений теоретически не существует для векторов, сконструированных на базе нитчатого бактериофага М13. Описаны случаи, когда в геном этого фага была встроена чужеродная ДНК длиной около 40 тыс. нуклеотидных остатков. Известно, однако, что фаг М13 становится нестабильным, когда длина чужеродной ДНК превышает 5 тыс. нуклеотидных остатков. Фактически же векторы, полученные из ДНК фага М13, используются главным образом для секвенирования и мутагенеза генов, и размеры встраиваемых в них фрагментов намного меньше.
Эти векторы конструируются из реплекативной (двутяжевой) формы ДНК фага М13, в которую встроены "полилинкерные" участки (пример такой конструкции показан на рис. 5). В фаговую частицу ДНК включается в виде однотяжевой молекулы. Таким образом, этот вектор позволяет получать клонированный ген или его фрагмент как в двутяжевой, так и в однотяжевой форме. Однотяжевые формы рекомбинантных ДНК широко используются в настоящее время при определении нуклеотидной последовательности ДНК методом Сэнгера и для олигодезоксинуклеотид-направленного мутагенеза генов.
Перенос чужеродных генов в клетки животных осуществляется с помощью векторов, полученных из ДНК ряда хорошо изученных вирусов животных - SV40, некоторых аденовирусов, вируса папиломы быка, вируса оспы и так далее. Конструирование этих векторов проводится по стандартной схеме: удаление "лишних" сайтов для рестриктаз, введение маркерных генов в области ДНК, не существенные для ее репликации (например, гена тимидин-киназы (tk) из HSV (вируса герпеса)), введение регуляторных районов, повышающих уровень экспрессии генов.
Удобными оказались так называемые "челночные векторы", способные реплицироваться как в клетках животных, так и в клетках бактерий. Их получают, сшивая друг с другом большие сегменты векторов животных и бактерий (например, SV40 и pBR322) так, чтобы районы, ответственные за репликацию ДНК, остались незатронутыми. Это позволяет проводить основные операции по конструированию вектора в бактериальной клетке (что технически намного проще), а затем полученную рекомбинантную ДНК использовать для клонирования генов в животной клетке.
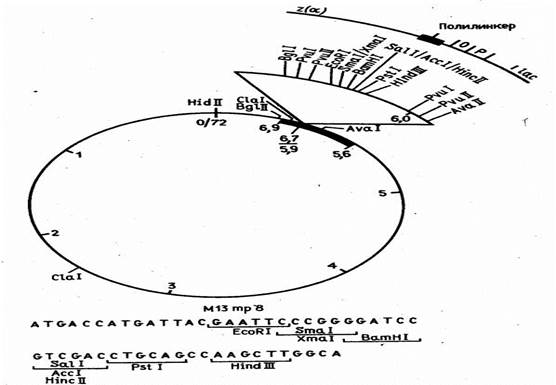
Рисунок 2. Рестрикционная карта вектора М13 mp8.








