 2020-01-14
2020-01-14 303
303
1) Відсутня лацентація.
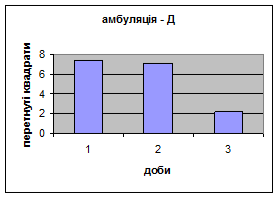
2) У самців Д-групи спостерігалась найнижча локомоція в порівнянні з тваринами інших груп. Це говорить про те, що світло для них був звичним фактором існування. На відміну від інших груп, серед переміщення даних тварин по “полю" було зареєстровано перетинання майже всіма піддослідними центра “поля”. Такий характер амбуляції не спостерігався серед інших експериментальних груп. Деякі Д-самці навіть тривалий час знаходились безпосередньо під лампою. Таким чином, на основі отриманих фактів, можна сказати, що тварини цієї групи майже не відчували страху бо дія світла для них була звичною. Показники амбуляції протягом 3х днів знижувалась з 7,38 до 2,19 (мал.12).
Малюнок 12
3) Вегетативні показники.
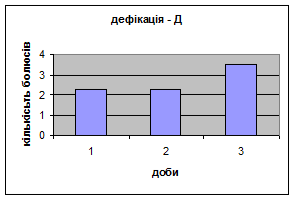
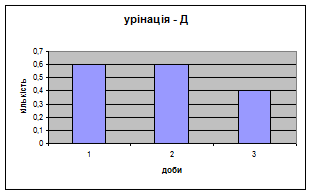
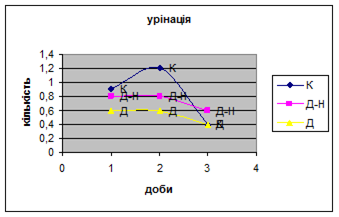
Показники дефікації зростали з 2,3 до 3,5 (мал.13), що говорить про швидке звикання до умов існування. Позитивна кореляція між показниками дефікації та локомоції свідчать також на цю користь. Найменша кількість урінацій (від0,6 до 0,4) (мал.14) була зареєстрована саме в цій групі самців, яка також мала позитивну кореляцію з амбуляцією.
Малюнок 12 Малюнок 13
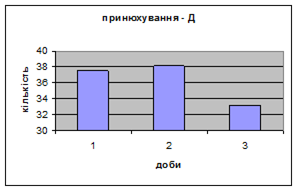
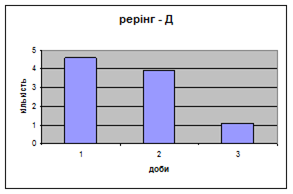
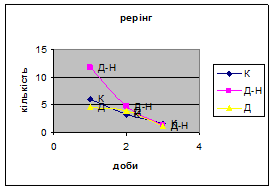
4) Самці Д-групи за показниками рерінгу були найменш активними серед експериментальних груп. Як і в попередніх групах дані показники цього параметру зменшувались (з 4,6 до 1,1) (мал.15). Але в порівнянні з К і Д-Н - групами, самці Д-групи протягом перших 3х днів експерименту проявляли стійки в напрямку до лампи, що не спостерігалось серед інших груп. Знову, це підтверджує думку про те, що дані тварини не відчували страху перед дією світла. Це, а також незначне зменшення кількості принюхувань (від 37,5 до 33,2) (мал14), дає можливість зробити висновок про швидкий розвиток у самців Д-групи звикання до стресових факторів середовища.
Малюнок 14 ёМалюнок 15
Висновки
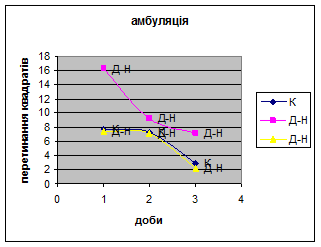
1. У всіх групах експериментальних тварин було зареєстровано зниження амбуляції (мал.16) і підвищення дефікації (мал.17).
Малюнок 16
Малюнок 17
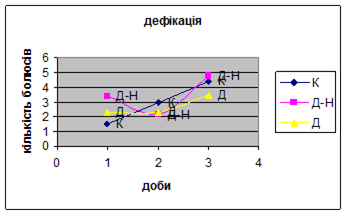
Але у самців Д-Н-групи відбувається більш повільніша редукція страху і емоційності, в порівнянні з іншими групами (К і Д) піддослідних тварин, так як, на другий день експерименту кількість болюсів у них різко зменшилась. У самців Д-групи спостерігалась найнижча локомоція, бо світло для них було звичним фактором існування. Також серед тварин цієї групи було зареєстровано перетинання майже всіма піддослідними центра “поля”, що не спостерігалось серед інших груп. Деякі Д-самці навіть тривалий час знаходились безпосередньо під лампою. Отже, тварини цієї групи майже не відчували страху, бо дія світла для них була звичною.
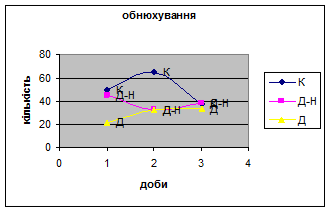
2. Загальне число принюхувань найбільше серед тварин Д-Н групи. На відміну від самців К - і Д - груп на другий день експерименту вони проявили найменшу нюхову активність - свій мінімум (мал.18), в той час як у останніх спостерігався її підйом, а далі поступове зниження.
Малюнок 18
3. Найбільший коефіцієнт активності (рерінг) був зареєстрований саме в Д-Н групі тварин (мал. 19). Отже, що дані самці більш важче пристосовуються до умов середовища, особливо в перші дні, де адаптація у них майже не спостерігається. Самці Д-групи за показниками рерінгу були найменш активними серед експериментальних груп. Але в порівнянні з К і Д-Н - групами, вони протягом перших 3х днів експерименту проявляли стійки в напрямку до лампи. Тобто, дані тварини не відчували страху перед дією світла.
Малюнок 19
4. Найменша кількість урінацій (від 0,6 до 0,4) (мал. 20) була зареєстрована саме в Д - групі самців. Загалом, у всіх групах вона поступово зменшується, що говорить про редукцію страху тварин.
Малюнок 20
5. Вище описані дані дають можливість стверджувати, що даним піддослідним самцям всіх груп властива емоційна, неагресивна, пасивно - оборонна поведінка (Кулагин Д.А., Федоров В.К., 1969). Тобто домінантними у поведінці даних тварин є стан психічної “відмови від пошуку”,“визнання поразки”. Як правило, наявність неспецифічних умов існування супроводжується появою психосоматичних захворювань. А це в свою чергу, при вище описаній формі поведінки може призвести до елімінації особин, перевага яких могла б негативно відбитись на життєдіяльності популяції вцілому, що сприяє еволюції виду і попереджає його регрес.
Отже, найповільніше адаптуються до стресових умов середовища самці Д-Н-групи, а найшвидше - самці Д-групи, так як вони підлягали цілодобової дії абіогенного фактора (світла) протягом 5 місяців, і за цей час встигли до нього звикнути. Про високу спроможність активно адаптуватись до стресових умов середовища свідчить і той факт, що у представників всіх груп був відсутній латентний період.
Література
1. Агаджанян Н.А. Биол. ритмы. - М.: Медицина., 1967. - 120с.
2. Ашофф Ю.А. Биол. ритмы: в 2х томах. Под ред. Ю.А. Ашоффа. - М.: Мир, Т.1. - 412с.
3. Биолог. ритмы. М.: Наука, 1980, 319с. Авт.: Ю.А. Романов,
С.А. Чепурнов, Г.А. Клевезоль.
4. Биологические часы. [Сб-к статей] Пер-д с англ. под ред. С.Э. Шноля. - М.: Мир, 1964. - 694с
5. Деряпа Н.Р. Проблемы мед. биоритмологии. - М.: Медицина., 1985. - 208с.
6. Дильман В.М. Большие биологические часы: Введение в интегральную медецину. - М.: Знание, 1981. - 208 с
7. Доскин В.А., Лаврентьева Н.А. Ритмы жизни. - 2-е изд. перераб. и доп. М.: Медицина, 1991. - 172с
8. Ефимов М.Л. Биол. ритмы в норме и патологии. - Алма-Ата. Казахстан, 1981. - 152с.
9. Зидерманс А.А. Некоторые вопросы хронобиологии и хрономедицыны: Обзор лит. / АН. Латв. ССР, Ин-т орган синтеза. Рига: Зинотне, 1988. - 212с
10. Лобашев М.Е. и Саватеев В.Б. Физиологические суточные ритмы животных.М. - Л. Изд-во Академии наук СССР, 1959. - 259с
11. Кенунен О.Г., Козловский В.Л. Структура двигательного поведения лабораторных животных - новые возможности методики “ Открытого поля”. // Физиолог. журн. им. Сеченова 1992 - Т.78, № 1
12. Комаров Ф.И., Рапопорт С.И. Хронобиология и хрономедецина. - М., “Триада-Х”, 2000. - 288с
13. Кузюта Э.И., Серохвостов А.П. Ритмы под солнечным ветром. - Фрунзе, Кыргыстан, 1991. - 104с.
14. Кулагин Д.А., Федоров В.К. Генетика поведения. - Ин-т физиологии им.А. Павлова. АН СССР. “ Наука”. 1969.
15. Маркель А.Л., Хусаинов Р.А. Метод комплексной регистрации поведенческих и вегетативных реакций у крыс при проведении теста “Открытого поля”. // Высшая нервная д-ть. 1976 - Т.26, №6.
16. Матюхин В.А. Биоритмология перемещений человека. - Новосибирск: “Наука”, Сиб. отделение, 1976. - 104 с
17. Морфофизиологический анализ формирования механизма цикла активность - покой в онтогенезе человека / А.И. Белич, М. Н. Константинова // Вестник Российской Академии мед. наук. - 1996 №3 ст55-61
18. Павлович Н.В. Биомагнитные ритмы. - Минск: Университетское, 1991. - 134с.
19. Путилов А.А. “Совы”, “Жаворонки” и др.: О наших внутренних часах и их влиянии на здоровье и характер. - Новосибирск: Изд-во Новосиб. ун-та.; М.: Совершенство, 1997. - 263с.
20. Ритмы жизни / Л.В. Львова. Провизор. - 2003 №1. - с.34-37
21. Руттенбург С.О. Циркадный ритм Физиологических процессов и трудовая деятельность человека. - Фрунзе: “Илим”, 1976. - 188с
22. Силантьева К.Г. Поведение и его механизмы: Учеб. пособие к спецкурсу [Поведение животных] / Урал. гос. пед. ин-т. - Екатеринбург: Урал. ГПИ, 1992. - 133
23. Уинфри А.Т. Время по биологическим часам / Пер. с анг. А.М. Алпатова; под ред.В.И. Кринского. - М.: Мир, 1990. - 208с
24. Фарби К.Э. Основы психозоологии: [Учеб. Длявузов по спец. “Психология”, “Биология”, “Зоология" и “Физиология”]. - 3-е изд. М.: Учеб. - метод. коллектор “Психология”, 2001. - 463.
25. Фельдман Г.Л. Биоритмология. Изд-во Ростовского ун-та, 1982. - 80с.
26. Хаснулин В.И. Космические тайны вашего самочувствия. Новосибирск: Наука. Сиб. отд-ние, 1992. - 176с.
27. Хронобиология и хрономедецина: Руков-во / Ф.И. Лимарова и др. под ред.Ф.И. Комарова. - М.: Медицина, 1989. - 399с
28. Hall C. S. 1934a Comp. Psychol., 17, p89.
29. Hall C. S. 1934b Comp. Psychol., 18, p385.
30. Hall C. S. and L. L. Klein. 1942.comp. Psychol., 33, p371.








